PODEM OS CRIACIONISTAS ACEITAR A ORIGEM DE
NOVAS ESPÉCIES?
Marcia Oliveira de Paula, Ph.D.
INTRODUÇÃO
No
capítulo 1 do livro de Gênesis é descrita a criação de todos os seres vivos por
Deus. Todos ele foram criados "segundo sua espécie". No início do
século dezenove, a maioria dos acadêmicos aceitava o relato de Gênesis bastante
literalmente. Muitos, mesmo, parece que iam além da declaração literal, e
afirmavam que as expressões encontradas em Gênesis 1 "segundo a sua
espécie" significavam que nenhuma variação poderia ocorrer dentro
das espécies básicas. Realmente, a Bíblia não fala em lugar algum de
"nenhuma variação". Este ensino relativo a nenhuma variação tem sido
chamado de Doutrina da Fixidez Extrema.
Darwin aparentemente iniciou sua
viagem ao redor do mundo crendo que "a ausência de variação"
constituía a verdade ensinada no livro de Gênesis. Porém, as sua observações, que revelaram muitas variações dentro
das espécies, puseram-no em confronto direto com o que entendia ser a doutrina
das origens expressa em Gênesis. Infelizmente, parece que Darwin não estudou
por si mesmo, com o devido cuidado, o livro de Gênesis. Acreditava que seus
mestres haviam interpretado Gênesis corretamente, e que a sua descoberta das
variações tinha mostrado que o relato de Gênesis não poderia ser aceito.
Tragicamente chegou à conclusão, como muitos desde então, que havia refutado o
livro de Gênesis, quando na realidade o que fez foi refutar uma de sua
interpretações extremadas. Quem ler Gênesis por si próprio nada encontrará dito
a respeito de ausência de variações. Descobrirá, sim, que os tipos básicos
foram criados conforme suas espécies.
Então, será que o criacionista pode
aceitar a origem de novas espécies? Alguns criacionistas fixistas acreditam que
Deus criou todas as espécies que existem atualmente. Mas esse conceito resulta
de uma má compreensão do termo "segundo a sua espécie" do livro de
Gênesis.
O
que são espécies? Segundo Mayr (1963) "As espécies são grupos naturais de
populações ativa ou potencialmente entrecruzantes e reprodutivamente isolados
de todos os outros grupos similares." Esta definição não se aplica,
entretanto, a todos os seres vivos. Certos organismos se reproduzem primária ou
exclusivamente por via assexual, o que torna inviável o teste do
entrecruzamento; e, algumas vezes, membros de diferentes espécies podem
entrecruzar-se com êxito.
Wolfe (1977) fornece uma definição de
espécie um pouco mais aberta: "A espécie é um grupo de organismos que se
assemelham entre si mais do que os organismos de um outro grupo qualquer e que
se distinguem dos integrantes de qualquer outro ao menos por uma característica
bem definida. Idealmente, os membros de uma espécie entrecruzam-se
exclusivamente entre si para produzir descendentes férteis."
As espécies de animais são
consideradas grupos naturais de populações que se cruzam efetivamente ou
potencialmente, mas as outras categorias da hierarquia taxonômica são
arbitrárias e estabelecidas pelo homem. Elas pretendem refletir níveis
diferentes de parentesco entre os animais, baseados na evolução. Duas ou mais
espécies com certos caracteres em comum formam um gênero. Por sua vez, gêneros
tendo caracteres comuns constituem uma família, as famílias são combinadas em
ordens, as ordens em classes e as classes em filos. Todos os filos juntos
constituem um reino.
Quais teriam sido então os tipos
básicos criados por Deus? Espécies, gêneros ou famílias? Seria bastante difícil
chegar a uma conclusão com os dados que temos, mesmo porque a classificação é
artificial, e, acima de espécie, não há critérios definidos. Mas é certo que
nem todas as espécies que existem foram criadas por Deus: muitas delas, e
provavelmente alguns gêneros também, surgiram por modificação dos tipos básicos
originais. Veremos a seguir alguns exemplos.
EXEMPLOS DE ESPECIAÇÃO
Existem vários exemplos atuais de
surgimento de novas espécies que foram documentados. No Havaí, existem diversas
espécies de mariposas do gênero Hedylepta
que se alimentam exclusivamente de bananeiras. Todas as outras espécies
havaianas desse gênero se alimentam de gramíneas, palmeiras ou leguminosas. O
fato notável neste caso é que as bananeiras foram introduzidas nas ilhas
do Havaí pelos polinésios há apenas mil
anos atrás. Aparentemente o gênero Hedylepta
sofreu eventos múltiplos de
especiação durante este breve período de tempo, que originaram espécies
que só se alimentavam das bananeiras recém-introduzidas no Havaí. Cada espécie
de mariposa que se alimenta de bananeiras está confinada às florestas de
montanhas de apenas uma ou duas ilhas. É compreensível esta distribuição das
espécies em questão: elas ainda não tiveram tempo de se espalharem para muito
longe de seus locais de origem.
Outro exemplo que pode ser mencionado
ocorreu na Uganda. O lago Nabugabo é um pequeno corpo de água doce desse país,
e fica situado às margens do lago Vitória. Evidências geológicas indicam que o
lago Nabugabo foi formado a partir do lago Vitória, pelo crescimento de um
banco de areia através de uma pequena enseada na margem deste lago. A datação
com radiocarbono indica que a separação ocorreu há aproximadamente quatro mil anos atrás. Um fato extraordinário
ocorre no lago Nabugabo: ele abriga quatro espécies de peixes da família dos
ciclídeos que não são conhecidas em nenhum outro lugar, inclusive no lago
Vitória, mas cada uma destas espécies se assemelha a uma espécie do lago
ancestral. As espécies do lago Nabugabo diferem das espécies parentais óbvias
do lago maior na coloração dos machos e em outras características menores. O
padrão de coloração do macho é uma importante característica na identificação
das espécies, servindo como um sinal de reconhecimento para o acasalamento. As
pequenas populações das espécies parentais do lago Vitória foram aparentemente
isoladas quando o banco de areia transformou o Nabugabo em um lago separado, e
desde então elas vieram a se tornar novas espécies.
Especiação por
poliploidia
Um mecanismo que pode levar à
especiação rápida é a poliploidia. Um indivíduo poliplóide é aquele que
apresenta um número múltiplo exato do genoma característico da espécie. Os
poliplóides são designados triplóides (3n), tetraplóides (4n), pentaplóides
(5n) etc., conforme tenham três, quatro, cinco ou mais genomas. Os poliplóides
podem se originar de duas maneiras principais:
a) por divisão
mitótica incompleta, havendo duplicação do número de cromossomos dentro da
própria espécie ou pela união de dois gametas não reduzidos (autopoliplidia)
b) por
hibridização interespecífica, seguida por divisão mitótica incompleta (alopoliploidia)
O geneticista russo Karpechenko foi o
primeiro a conseguir obter uma espécie nova em laboratório usando a
alopoliploidia. Ele cruzou rabanete, Raphanus
(2n =18), com couve, Brassica (2n = 18), desejando obter
descendentes com raiz de rabanete e folhas de couve. Para sua decepção, o que
conseguiu foram plantas com raiz de couve e folha de rabanete. O híbrido obtido
era estéril, pois seus 18 cromossomos não se pareavam na meiose, sendo por isso
distribuídos irregularmente nos gametas. A duplicação do número de cromossomos
nos híbridos produziu plantas tetraplóides que se mostravam férteis, pois nelas
9 cromossomos da couve pareavam com 9 da couve, e 9 do rabanete com 9 do
rabanete. Este tetraplóide recebeu o nome de Raphanobrassica e mostrou-se uma nova espécie, geneticamente
isolada, tanto da couve como do rabanete, infelizmente sem as qualidades
apreciadas daquelas plantas.
Os alopoliplóides, sendo geneticamente
isolados dos tipos parentais, constituem novas espécies, nas quais estão
combinadas muitas das características de seus ancestrais.
Novas espécies alopoliplóides podem
também ser formadas na natureza. Um dos poliplóides mais bem conhecido é uma
gramínea de brejo salgado do gênero Spartina.
Uma espécie nativa deste gênero, S.
maritima, ocorre em brejos ao longo da costa da Europa e da África. Uma
segunda espécie, S. alterniflora, do
leste da América do Norte, foi acidentalmente introduzida na Grã-Bretanha por
volta do ano de 1800 e desenvolveu-se formando grandes colônias locais. A
espécie nativa S. maritima possui
estatura baixa, mas S. alterniflora é muito mais alta,
alcançando freqüentemente 0,5 metro de altura e, ocasionalmente, um metro. Nas
proximidades do porto em Southampton, tanto a espécie nativa quanto a
introduzida existiram lado a lado , e, em 1870, coletou-se um híbrido estéril,
que se reproduzia por rizomas. S.
maritima possui um número somático de cromossomos (2n) de 60 e S. alterniflora, 62; o híbrido, devido a
alguma divisão meiótica errada, possui também 62 cromossomos somáticos. Este
híbrido estéril, que foi denominado S.
townsendii persiste até hoje. Por volta de 1890, um vigoroso poliplóide
fértil originou-se naturalmente deste híbrido espalhando-se rapidamente por
todo o litoral da Grã-Bretanha e noroeste da França. Este poliplóide, S. anglica, com 122 cromossomos, surgiu
devido à duplicação dos cromossomos do híbrido. S. anglica é freqüentemente plantado para ligar o lodo dos brejos e
tal uso contribuiu para sua rápida disseminação. Na atualidade, está substituindo
ambas as espécies parentais.
O PROCESSO DE ESPECIAÇÃO
O
material genético de uma população, interagindo com o meio e submetido às
forças da mutação, seleção natural, deriva genética e migração, pode levar ao
aparecimento de divergência dentro dessa população. Diferenças substanciais
podem ocorrer entre dois grupos de uma dada população, a ponto de se poder
identificá-los como entidades distintas. Essas diferenças freqüentemente estão associadas
à existência de regiões ecologicamente distintas, fato que leva esses grupos a
se adaptarem de maneira peculiar. Caracteres de alto valor adaptativo em uma
região podem se comportar de maneira diferente em outras. As constituições
genéticas de dois ou mais setores de uma população são passíveis de diversificação, por estarem submetidos a pressões
seletivas distintas. Se do cruzamento entre membros de dois grupos resultar
descendência fértil, admite-se que esses dois grupos constituam raças. As raças
são definidas como populações da mesma espécie, que diferem nas freqüências
relativas de genes ou de formas cromossômicas.
Se os mecanismos de isolamento
tornarem-se cada vez mais eficientes e o fluxo gênico (intercâmbio gênico
através da reprodução) entre as raças for cada vez menor, elas tenderão a
divergir até o ponto em que a reprodução entre elas se torne impossível. Quando
isto ocorrer, o processo de diversificação tornar-se-á irreversível, não haverá
mais troca de genes entre os dois grupos e estes poderão, agora, ser
considerados duas espécies distintas (Ver Figura 1).
Este
processo de especiação descrito por Stebbins pode ser perfeitamente aceito
pelos criacionistas.
O tempo requerido para a formação de
uma nova espécie por este processo seria, segundo alguns evolucionistas, em
média um milhão de anos. Porém, vários outros cientistas admitem que este
processo possa ser muito mais rápido, sendo possível em centenas ou milhares de
anos (ver exemplos acima).
Segundo
Gibson, o dilúvio e as condições originadas após o seu término teriam fornecido
condições muito favoráveis para uma rápida especiação. A maioria dos organismos
foi destruída por esta catástrofe, deixando pequenas populações de
sobreviventes . Os vertebrados terrestres foram preservados na arca em pequenos
números. Após eles serem liberados da arca, eles teriam encontrado recursos
quase ilimitados disponíveis, tornando possíveis rápidos aumentos no tamanho
das populações, juntamente com níveis reduzidos de competição. Haveria um
grande número de nichos ecológicos desocupados, aos quais os organismos
poderiam se adaptar. As condições
ambientais da terra estariam instáveis, e processos geológicos como os vulcões,
terremotos, e mudanças no nível do mar afetariam o clima, criariam e removeriam
barreiras para a dispersão e produziriam muitas catástrofes localizadas que
tenderiam a isolar populações de espécies em dispersão.
Aquelas espécies que foram preservadas
fora da arca também estariam sujeitas a condições favoráveis para especiação.
Organismos aquáticos poderiam ser transportados por correntes, possivelmente
resultando na dispersão de pequenos grupos de sobreviventes para muitos lugares
isolados com diferentes condições ambientais. O mesmo poderia acontecer com
grupos terrestre tais como insetos, vermes e outros invertebrados. Plantas e
sementes poderiam também ser levadas pelas águas e dispersas por correntes.
Estas condições provavelmente resultariam em especiação rápida em muitos grupos
de organismos.
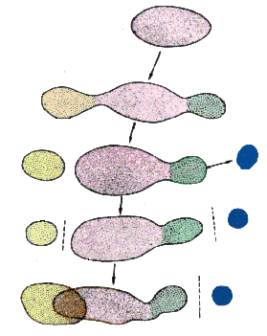
1o estágio
Uma única população em
um ambiente homogêneo.
2o estágio
Diferenciação do
ambiente e migração para novos ambientes produzem diferenciação.
3o estágio
Modificações
posteriores e migrações conduzem ao isolamento geográfico de algumas raças e
subespécies.
4o estágio
Algumas dessas subespécies isoladas se diferenciam no tocante a modificações gênicas e cromossômicas que controlam mecanismos de isolamento
5o estágio
Modificações no ambiente permitem
que populações geograficamente isoladas coexistam novamente na mesma região.
Elas agora permanecem distintas por causa das barreiras de isolamento
reprodutivo que as separam e podem ser reconhecidas como espécies distintas.
Fig. 1 -
Diagrama mostrando a seqüência de acontecimentos que leva à diferenciação de
raças, subespécies e espécies, tendo como ponto de partida um grupo homogêneo
de populações (segundo Stebbins, 1974).
CONCLUSÃO
Mudança
nas espécies através dos tempos são irrefutáveis. Porém, essas mudanças são
limitadas. A possibilidade de mudanças nas espécies não deveria surpreender os
criacionistas. A má compreensão do termo "segundo sua espécie" no
livro de Gênesis levou alguns a pensarem que os animais não podem mudar de
maneira significativa. Porém uma leitura cuidadosa mostra que o texto está
afirmando que Deus criou muitos tipos de organismos em um dia de criação. O
termo não diz nada sobre se eles podem ou não mudar. Ao contrário, o livro de
Gênesis claramente afirma que mudanças iriam acontecer (Gênesis 3: 14, 18).
Parece então lógico aceitar o conceito de que Deus criou os "tipos"
básicos de organismos, originando a
grande variedade de vida ao nosso redor, mas ocorreram mudanças morfológicas
limitadas e formação de novas espécies e talvez gêneros. Estas mudanças podem
ter acontecido em tempos relativamente curtos, após a criação.
BIBLIOGRAFIA
BRAND, L. R.
& GIBSON, J. An interventionist theory of natural selection and
biological change within limits. Origins,
2: 60 - 82, 1993.
CARVALHO, H.C. Fundamentos
de Genética e Evolução. 3a ed. Rio de Janeiro, Livraria Atheneu,
1987. 556 p.
GARDNER, E. J., SIMMONS, M. J. & SNUSTAD, D. P. Principles
of Genetics. 8a ed. New York, John Wiley & Sons, 1991. 649
p.
GIBSON, L. J.
Are there limits to change in species? Geoscience
Research Institute
MARSH, F. L.
Variação e fixidez entre os seres vivos - um novo princípio biológico. Folha Criacionista, 21: 17 - 24, 1979.
MAYR, E. Animal species and evolution.
Cambridge, Mass., Belknap Press, Harvard Univers. Press,
1963. 797 p.
MORRIS, H. M. O
enigma das origens: a resposta. Belo Horizonte, Editora Origens, 1995. 265
p.
RAVEN, P. H.,
EVERT, R. F. & CURTIS, H. Biologia vegetal. 2a ed. Rio de
Janeiro, Editora Guanabara, 1978. 724 p.
STANLEY, S. M. The new evolutionary timetable. Fossils,
genes and the origin of new species.
New York, Basic Books Inc. Publishers,
1981.
STEBBINS, G. L. Processos
de evolução orgânica. São Paulo, Editora da Universidade de São Paulo,
1970. 255 p.
WEBSTER, C. L. A
perspectiva de um cientista sobre a criação e o dilúvio. São Paulo, Editora
Universitária Adventista, 1999. 31 p.
WOLFE, S. L. Biology: the foundations. Belmont,
Califórnia, Wadsworth Publishing Company, 1977. 460 p.