
Translated by Dr. Ubaldo Leme Filho
INTRODUCTION
The first chapter of the book of Genesis describes the creation of all living beings by God. All of them were created "according to their species". In the beginning of the nineteenth century, most scholars accepted the report of Genesis quite literally. Many, it seems went beyond what was stated literally, and affirmed that the expressions found in Genesis 1 "according to his/her species" meant that no variation could happen within the basic species. Really, the Bible doesn't speak anywhere of any "variation" that took place. This relative teaching the no variation has been called the "Doctrine of Extreme Fixation".
Darwin seemingly began his trip around of the world believing that the "absence of variation" constituted the truth taught in the book of Genesis. However, his observations, that revealed a lot of variations inside of the species, put him in direct confrontation with what he understood to be the doctrine of the origins stated in Genesis. Unfortunately, it seems that Darwin didn't study by himself, with due care, the book of Genesis. He believed that his masters had interpreted Genesis correctly, and that his findings of the variations had shown that the report of Gênesis could not be accepted. Tragically he reached the conclusion, as many since then, that it had refuted the book of Genesis, when in reality what he did was to discredit one of its more extreme interpretations. Whoever reads Genesis will find nothing stated regarding absence of variations. He will discover, however, that the basic types were created according to their species.
Can we then conclude that a criacionist can accept the origin of new species? Some "fixist" creationists believe that God created all of the species that exist now. But that concept results from a misunderstanding of the term "according to their species" in the book of Genesis.
What are species? According to Mayr (1963) "Species are naturally occurring active groups of populations or potentially cross breeding and reproductively isolated of all other similar groups." This definition does not apply, however, to all living beings. Certain organisms reproduce primary or exclusively through asexual mode, which makes it impossible to test cross breeding capability; and, sometimes, members of different species can interbreed successfully.
Wolfe (1977) supplies a definition of species that is a little more open: "Species is a group of organisms that resemble each other amongst themselves more than the organisms of any other group and that differ from members of any other in at least one well defined trait or characteristic. Ideally, the members of a species breed exclusively amongst themselves to produce fertile offsprings."
Species of animals are considered natural groups of populations that breed effectively or potentially, but the other categories of the taxonomic hierarchy are arbitrary and established by humans. They intend to reflect different levels of relationship among the animals, based on evolution. Two or more species with certain characters in common form a gender. In its turn, genders having common traits or characters constitute a family, the families are combined in orders, the orders in classes and the classes in phylum. All of the phyla together constitute a kingdom.
Which ones would have been the basic types created by God? Species, genders or families? It would be quite difficult to arrive at a conclusion with the data that we have, especially because the classification is artificial, and, above species, there are no defined criteria. But it is accepted that not all of the species that exist were created by God: many of them, and probably some genders also, appeared from modifications of the original basic types. We will show some examples below.
EXAMPLES OF SPECIATION
Various current examples of appearance of new species have been documented. In Hawaii, there are several species of moths of the Hedylepta gender that feed exclusively on banana trees. All the other Hawaiian species of that gender feed on grass, palm trees or legume. The notable fact in this case is that the banana trees were introduced at the islands of Hawaii by the Polynesians just a thousand years ago. Seemingly the gender Hedylepta suffered multiple events of speciation during this brief period of time, that originated species that only fed on the banana trees "recently" introduced in Hawaii. Each moth species that feeds on banana trees is confined to the forests on mountains of just one or two islands. This distribution of these species here discussed is understandable: they still didn't have time to spread very far away from their places of origin.
Another example that can be mentioned happened in Uganda. Lake Nabugabo is a small body of fresh water in that country, and it is located on the margins of lake Victoria. Geological evidences indicate that the lake Nabugabo was formed from lake Victoria, by the growth of a sandbank through a small cove in the edge of this lake. The dating with radiocarbon indicates that the separation happened approximately four thousand years ago. An extraordinary fact occurs in lake Nabugabo: it shelters four species of fish of the ciclideos family that are not known to exist in any other place, including lake Victoria, but each one of these species resembles other types or species from the ancestral lake. The species of lake Nabugabo differ from the obvious parental species of the larger lake in the coloration of the males and in other minor characteristics. The pattern of coloration of the male is an important characteristic in the identification of the species, serving as a sign of recognition for mating purposes. Small populations of the parental species on lake Victoria were apparently isolated when the sandbank transformed Nabugabo in a separate lake, and since then, they have become new species.
Speciation by polyploidy
A mechanism that can allow for fast speciation is polyploidy. A polyploid organism is one that presents an exact multiple number of the genomic characteristics for a species. The polyploid organisms are designated triploids (3n), tetraploids (4n), pentaploids (5n) etc., as they have three, four, five or more genomes. The polyploids can originate mainly by two different paths or mechanisms:
- by incomplete mitotic division, having duplication of the number of chromosomes within the species or by the union of two non reduced gametes (autopolyploidy)
- by interspecies hybridization, followed by incomplete mitotic division (allopolyploidy)
The Russian geneticist Karpechenko was the first to obtain a new species in laboratory using the allopolyploidy. He crossed radish, Raphanus (2n =18), with collard greens, Brassica (2n = 18), trying to obtain descendents with radish root and collard greens leaves. For his diappointment, what he got were plants with collard greens root and radish leaf. The hybrid specimens obtained were sterile, because their 18 chromosomes did not pair in meiosis, being distributed irregularly in the gametes. The duplication of the number of chromosomes in the hybrid plants produced tetraploid specimens that were shown to be fertile, because in them 9 chromosomes of the collard greens paired with 9 of the collard greens, and 9 of the radish with 9 of the radish. This tetraplóide received the name of Raphanobrassica and became a new species, genetically isolated, boh from the collard greens and from the radish, unfortunately without the appreciated qualities of those plants.
The allopolyploids, being isolated genetically of the parental types, constitute a new species, in which many of the characteristics of their ancestral ones are combined.
New allopolyploid species can also be formed in nature. One of the polyploids better known is a gramineous of salty swamps of the gender Spartina. A native species of this gender, S. maritima, occurs at swamps along the coasts of Europe and Africa. A second species, S. alterniflora, from eastern North America, was introduced accidentally in Grã-Britain around the year of 1800 and it grew forming large local colonies. The native species S. maritima has low stature, but S. alterniflora is much taller, frequently reaching 0,5 meter of height and, occasionally one meter. In the vacinities of the port in Southampton, both the native species and the one introduced coexisted side by side, and, in 1870, a sterile hybrid was collected, that reproduced by rhizomes. S. maritima possesses a somatic number of chromosomes (2n) of 60 and S. alterniflora, 62; the hybrid, due to some erratic mitotic division, also possesses 62 somatic chromosomes. This sterile hybrid, S. townsendii as it came to be called persists until today. Around 1890, a vigorous fertile poliplóide originated naturally from this hybrid one dispersing quickly through the whole coast of Great-Britain and the northwest of France. This polyploid, called S. anglica, with 122 chromosomes, appeared due to the duplication of the chromosomes in the hybrid. S. anglica is frequently planted to fix the mud of the swamps and such use contributed to its fast dissemination. At the present time, it is substituting both parental species.
THE PROCESS OF SPECIATION
The genetic material of a population, interacting with the environment and submitted to the forces of mutation, natural selection, genetic derivations and migration, can lead to the emergence of divergences in that population. Substantial differences can happen among two groups of a given population, to the point of identifying them as different entities. Those differences frequently are associated with the existence of ecologically distinct areas, a fact that forces those groups to adapt in a peculiar way. Characteristics of high adaptative value in an area can behave in a different way in others. The genetic constitutions of two or more distinct sections of a population are susceptible to diversification, since they are normally submitted to different selective pressures. If the cross breeding among members of two different groups results in fertile offsprings, conceptually it is admitted that those two groups constitute races. Races are defined as populations of the same species, that differ in the relative frequencies of genes or chromosome forms.
If the mechanisms of isolation become increasingly more efficient and the genetic flow (exchange of genes through reproduction) between the races decreases correspondingly becoming ever smaller, they will tend to diverge up to the point in which reproduction between them becomes impossible. When this happens, the diversification process will become irreversible, there won't any be more exchange of genes between the two groups and these will be able to, now, be considered two different species (See Illustration 1).
This speciation process described by Stebbins can be accepted perfectly by the creationists. The time required for the formation of a new species by this process would be, according to some evolutionists, around a million years on the average. However, several other scientists admit that this process can be much faster, being possible in hundreds or thousands of years (see examples above).
According to Gibson, the flood and the conditions originated after it ended would have supplied very favorable conditions for a fast speciation. Most of the organisms were destroyed by this catastrophe, leaving small populations of survivors. The terrestrial vertebrates were preserved in the ark in small numbers. After they left the ark, they would have found almost limitless resources available, favoring rapid increases in the size of the populations, along with reduced levels of competition. There would be a great number of unoccupied ecological niches, to which the organisms could adapt. The environmental conditions of the earth would be unstable, and geological processes such as volcanic action, earthquakes, and changes in sea level would affect the climate, creating and removing barriers for dispersion at a fast pace, and producing many localized catastrophes that would tend to isolate populations of species in dispersion.
Those species that were preserved out of the ark would also be subject to favorable conditions for speciation. Aquatic organisms could be transported by currents, possibly resulting in the dispersion of small groups of survivors at many isolated places with different environmental conditions. The same could happen with terrestrial groups such as insects, worms and other invertebrates. Plants and seeds could also be taken by the waters and dispersed by sea or wind currents. These conditions would probably result in fast speciation in many groups of organisms.
Stages for Speciation
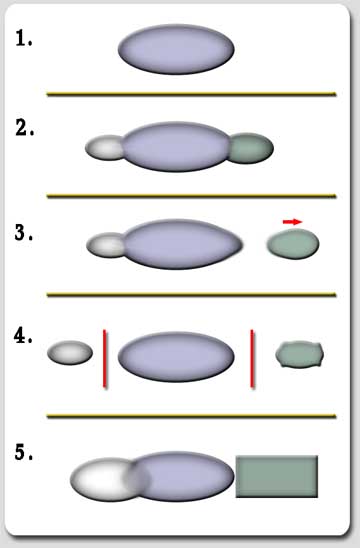
1. A single population in a homogeneous atmosphere.
2. Differentiation in the environment and migration to new surroundings produce differentiation.
3. Subsequent modifications and migrations lead to the geographical isolation of some races and subspecies.
4. Some of those isolated subspecies differentiate in areas such as genetic modifications and chromosome changes that control isolation mechanisms.
5. Modifications in the atmosphere allow for geographically isolated populations to coexist again in the same area. They now remain different because of the barriers of reproductive isolation that now separate them and they can be recognized as different species.
Fig. 1 - diagram showing the sequence of events that cause the differentiation of races, subspecies and species, having as a starting point a homogeneous group of populations (according to Stebbins, 1974).
CONCLUSION
Change in the species through the times is irrefutable. However, those changes are limited. The possibility of changes in the species should not surprise the creationists. The misunderstanding of the term"according to their species" in the book of Genesis led some to think that the animals cannot change in a significant way. However a careful reading shows that the text is affirming that God created many types of organisms in certain days of creation. The term doesn't say anything on whether they can or can not change. On the contrary, the book of Genesis clearly affirms that changes would happen (Genesis 3: 14, 18). it seems then, logical to accept the concept that God created the basic "types" of organisms, originating the great variety of life around us, but limited morphologic changes occured with formation of new species and eventually genders. These changes may have happened in a relatively short time following creation.
REFERENCES
BRAND, L. R. & GIBSON, J. An interventionist theory of natural selection and biological change within limits. Origins, 2: 60 - 82, 1993.
CARVALHO, H.C. Fundamentos de Genética e Evolução. 3a ed. Rio de Janeiro, Livraria Atheneu, 1987. 556 p.
GARDNER, E. J., SIMMONS, M. J. & SNUSTAD, D. P. Principles of Genetics. 8a ed. New York, John Wiley & Sons, 1991. 649 p.
GIBSON, L. J. Are there limits to change in species? Geoscience Research Institute
MARSH, F. L. Variação e fixidez entre os seres vivos - um novo princípio biológico. Folha Criacionista, 21: 17 - 24, 1979.
MAYR, E. Animal species and evolution. Cambridge, Mass., Belknap Press, Harvard Univers. Press, 1963. 797 p.
MORRIS, H. M. O enigma das origens: a resposta. Belo Horizonte, Editora Origens, 1995. 265 p.
RAVEN, P. H., EVERT, R. F. & CURTIS, H. Biologia vegetal. 2a ed. Rio de Janeiro, Editora Guanabara, 1978. 724 p.
STANLEY, S. M. The new evolutionary timetable. Fossils, genes and the origin of new species. New York, Basic Books Inc. Publishers, 1981.
STEBBINS, G. L. Processos de evolução orgânica. São Paulo, Editora da Universidade de São Paulo, 1970. 255 p.
WEBSTER, C. L. A perspectiva de um cientista sobre a criação e o dilúvio. São Paulo, Editora Universitária Adventista, 1999. 31 p.
WOLFE, S. L. Biology: the foundations. Belmont, Califórnia, Wadsworth Publishing Company, 1977. 460 p.