 |
Origen Abiogenica de la Vida: Una Teoria en Crisis
Arthur V. Chadwick, Ph. D.
Professor of Geology and Biology
Southwestern Adventist College
Keene, TX 76059 U.S.A.
El estudio del Origen de la Vida Como Una Ciencia:
Todos los fenomenos son únicos e irreproducibles. Es el propósito del método científico es relacionar el efecto (observación) con la causa al tratar de reproducir el efecto, recreando las condiciones bajo las cuales este ocurrió. Cuanto más complejo es el fenómeno, mayor es la dificultad que la ciencia tiene para investigarlo. En el caso de la investigación científica en relación con el origen de la vida, encontramos dos problemas: Las condiciones en que ocurrió son desconocidas y el fenómeno (la vida) tan complejo que nosotros no entendemos sus propiedades esenciales. Así mismo existe una condición de "desconocimiento" sobre el origen de la vida, muy diferente con aquel asociado con la mayoría de las investigaciones científicas. Una vez que los métodos científicos y el rigor de las investigaciones científicas están en desventaja en este punto, debemos estar más propensos a considerar toda información de cualquier fuente que pueda contribuir para un mayor entendimiento sobre los orígenes, en especial la hipótesis de que la vida fué creada por un Ser infinito y superior. Si un análisis científico cuidadoso nos lleva a concluir que los mecánismos propuestos del origen espontáneo no pudiera haber producido una célula viva, y también que el hecho de que ningún proceso natural inmaginativo pudiera haber resultado en el origen espontáneo de la vida, la hipótesis alternativa de la creación se torna más atrayente. Si por otro lado, consideramos que los mecanismos propuestos son plausibles, debemos estar concientes de que los métodos de la ciencia nunca podrían responder con certeza a la pregunta del origen de la vida. Vamos a comenzar a hacer consideraciones sobre la naturaleza de la vida y despues trataremos de evaluar el estado actual de la investigación sobre el origen de la vida, también conocida como "Paleobiogeoquímica".
¿Que es Vida?
Las celulas son los elementos más simple de todos los seres vivos. Todos los organismos complejos estan compuestos por células. Las celulas también son estructuras muy complejas. Dentro de cada célula existe un microcosmo de organismos del cual es una parte funcional. Células unicas de vida libre, como la ameba y otros protozarios, deben desempeñar todas las funciones necesarias para la vida, como los organismos más complejos. Estas células poseen un sistema digestivo, reprodutor, repiratorio, nervioso, esquelético, excretor, muscular, etc., minúsculos y muy complejos. Ademas, todas las células vivas poseen características en común que son consideradas fundamentales para la vida. Estas incluyen: un código genético, el código primario rico en informaciones de ADN, una enzima de ADN polimerasa que es necesaria para la reproducción del código, ARN intermediarios y el ARN polimerasa requerido para transcribir el ADN, un mecanismo de síntesis de proteínas envolviendo los ribosomas, los ARN transportadores y las enzimas necesarias para ligar a cada una de los aminoácidos respectivos, una menbrama celular y las vias metabólicas fundamentales para generar los materiales necesarios para las reacciones mencionadas anteriormente, envolviendo cientos de enzimas. Tal vez la primera célula viva no tenía la capacidad de realizar las actvidades más simples de una célula actual. En este caso, debemos decidir que características podemos eliminar, y despues de eliminarlas, verificar si la célula puede sobrevivir. Después, debemos explicar como estas caracteristicas surgieron en el proceso de evolución y por qué son comunes a todas las células. Discutiremos más sobre esas características más adelante. Ahora, vamos a dar una ojeada al sistema propuesto para el origen de la célula.
¿Como Piensan los Autores de la Teoría Abogenica de la Vida?
Fueron dados cuatro componentes escenciales para el origen de la vida:
1. Una atmósfera llena de moléculas gaseosas reducidas y una fuente de energía para convertir esas moléculas en precursores biológicos necesarios para la vida.
2. Un océano lleno de pequeñas moléculas biológicas resultantes del punto anterior.
3. Un mecanismo que genera, a partir de este océano de moléculas , polímeros ricos en información necesarios para una célula viva (ADN,ARN y proteínas)
4. Una crencia que si el paso número tres es implementado, resulltará casi inevitablemente, en la formación de una célula viva.
Examinaremos cada una de estas sequencia:
Evolución Química:
Los primeros estudios serios sobre el origen de la vida datan de 1920, cuando J.B.S. Haldane y A.I. Oparin sugirieron que la vida se originó espontaneamente de la materia no viva de la superficie de la tierra en un pasado remoto, y describieron un escenario para esa ocurrencia. Desde entonces una vista que se tenía era que la vida no era mas que química complicada, sus ideas se tornaron ampliamente aceptadas entre aquellos que procuraban establecer un origen naturalista para la vida en la tierra. Más no fué hasta el año 1953 que Stanley Miller hizo sus ahora famosos experimentos usando el reducido ambiente presentado por Oparin (CH3, NH4, H2O, and H2 ) en un vaso de vidrio energizado por un resorte de Tesla, que consiguió crear varios compouestos simples, incluyendo algunos aminoácidos, así como una cantidad de "alquitrán" (sedimento orgánico polimerizado, de poco interes para los paleobiogeoquímicos.) Miller y Urey propusieran que la luz ultravioleta (UV), descargas eléctricas y relámpagos producieron pequeñas moléculas biológicas precursoras en la "tierra primitiva", que subsequentemente fueron depositadas en los océanos por el ciclo hidrológico. Carl Sagan propuso que la tierra primitiva estaba sometida a un flujo de rayos UV 100 veces más fuerte que lo de hoy día, y que el H2S proveniente del vulcanismo era un agente que catalizava la transferencia de energía de la luz UV para los elementos transparentes UV de la atmósfera. En el inicio de la decada de los 70, Bar-Nun demostró que las ondas de choque de alta velocidad eran 10,000 veces tan eficientes en cuanto los otros métodos, en convertir la atmósfera gaseosa reductora de Oparin en pequeñas moléculas, así mismo formando cuatro aminoácidos. En los años más recientes, bajo condiciones un poco sospechosas, Yuasa (1984) consiguió producir purinas y pirimidinas (tipos de bases presentes en DNA y en RNA.) Actualmente, 14 de los 20 aminoácidos pueden ser producidos bajo condiciones de la atmósfera reductora que se creen que existió en la tierra primitiva. Desafortunadamente para los que acreditan por el origen espontánea de la vida, la mayoría de los aminoácidos producidos por estos experimentos son glicerina or alanina. Asi mismo, varios aminoacidos no proteicos son producidos, lo que compite con los veinte aminoacidos proteicos en cualquier reacción abiogénica.
Existen muchos problemas más, para aquellos que quieren acreditar el origen de vida espontánea, la articulación de un modelo para producir la "sopa diluida caliente" de Haldane y Oprain o la "sopa diluida de gallina" de Sagan, no importa cuan insatisfactoria pueda ser, nos acredita que estos modelos pueden explicar el origen de la vida. Vamos a caminar a través de sistemas modernos del estudió de la vida y vamos a tratar de evaluar estas especulaciones dentro de los parámetros que los investigadores han establecido para alcanzar la meta propuesta, ie. el origen espontáneo de una célula viva. Vamos a comenzar por considerar la atmósfera primitiva de la tierra y la probabilidad que esta pueda haber generado un océano lleno de moléculas muy útiles. Después vamos a considerar si podemos producir todas clases de polímeros biologicamente importantes, necesarios para la vida, dado un océano lleno de pequeñas moléculas. Después nos preguntaremos si es posible hacer una célula viva y investigar algunas áreas muy significativas de la biología molecular para identificar la complejidad que un sistema vivo abarca.
Evidencia para una atmósfera en Reducción:
Oparin sugirió una atmósfera para la "tierra primitiva" de H2, H2O, NH3, CH4 en su mayoría debido a que él, siendo un químico, reconoció que semejante atmósfera reducida iba a requerir, químicamente, la producción de una "sopa caliente diluida" de la que el creia que la vida se originaba. Siendo un químico, el también reconoció la necesidad de excluir oxigeno o algunos otros compuestos oxidativos de la mezcla. Esto fué muy conveniente, debido a que tal mezcla probó ser capaz de generar una variedad de pequeñas moléculas de interes biológico. La verdadera pregunta es, ¿Existió tal atmósfera en la tierra? Un analisis cuidadoso desde el punto de vista geológico, cosmológico, y químico reveló que semejante atmósfera de reducción, si de veras existió, no debio haber durado por mucho tiempo. J. C. G. Walker dice que " La más grande evidencia [de una atmósfera de reducción] es provista por las condiciones [requeridas] para el origen de la vida. Una atmósfera reducida es requerida."(Walker, 1976). Philip Abelson (1966) y J. W. Schopf (1972) concluyeron de que no hay evidencias para la existencia de una atmósfera de metano-ammonia. Desde el vuelo del Apollo 16, nos hemos dado cuenta que la fotodisociación del agua en la atmósfera superior por la inducción es una de las fuentes mayores de oxígeno libre atmosférico. Tal oxígeno habría sido producido en un promedio alto en la tierra primitiva, sin la presencia de un escudo de ozono (hecho por oxígeno) para bloquear la intensa luz UV del sol. Un analisis de las rocas sedimentarias primitivas del Precambrico parece indicar la presencia de oxígeno libre, quizas a los niveles similares de los hoy día (Walker, 1977). Mientras que se conoce con seguridad de las condiciones requeridas para el desarrollo de formaciones con bandas de hierro, rocas con cantidades significantes de oxidos ferrosos, podemos decir con seguridad que oxígeno libre se encontraba presente. Por muchos años estas formaciones fueron consideradas como el fenómeno del Proterozoico, y en su aparición simultanea con la aparición de una atmosfera oxidativa de 2.0 billones de años. Ahora nosotros conocemos que las formaciones con bandas de hierro y otros sedimentos oxidados, y por consiguiente que oxigeno libre estuvo presente a través del Archean (Schidlowski, 1976) y que las evidencias más primitivas de formas probables de vidas (de 3.7 billones de años) no es más primitivo que las formaciones más primitivas de bandas de hierro.
Otros datos en contra de una atmósfera de reducción han sido acumulados en los últimos veinte años. Muchos que en el pasado cosideraron una atmósfera reducida como único requisito, estan tomando un segundo vistaso. Muchas consideraciones teóricas requieren que la atmósfera se ha prodicido por la outgassing of the mantle, y cuyos gases hoy se han oxidado uniformemente. La probabilidad de una atmósfera neutra(CO2, H2O, N2, y posiblemente de una pequeña cantidad de H2) ha sido aprovada por muchos investigadores de esta área. Este prospecto no parece haber apagado con el entusiasmo de muchos trabajadores apreciablemente. Sin embargo la presencia de oxigeno libre entorpece todos los escenarios propuestos por el abiogenensis de las formas de vida, y tal atmósfera aparece con una seguridad virtual.
Evidencia para la "Sopa de Pollo":
Un número de analises cuidadosos del escenario de Oparin-Haldane para producir un océano lleno de pequeñas moleculas precursoras biologicas, ha dejado vacios en el concepto de un océano de "sopa diluida" en la tierra primitiva. Cuando el lapiz es puesto sobre el papel para calcular cuantas moléculas pudieron resultar bajo las condiciones ideales, la probabilidad de semejante océano se desvanace. H. E. Hull (1960), L. G. Sillen (1965) y R. Shapiro (1986) han concluido de que el término "diluido" es una enorme exageración y que la presencia de aún más abundantes aminoácidos no pudo haber excedido .0001 gramos/litros, demasiado diluido para haber estado envuelto en una reacción polimerica requerida para hacer proteinas. H. R. Hulett (1969) piensa que .000001 g/l es una cantidad más real para glycine, el amino ácido más abundante, y que K. Dose sugirió que esa concentración sería de .00001 g/l. Concentraciones presentes en el Atlántico mediano oscila entre .00001 y .0001 g/l! Si la síntesis de pequeñas moléculas de la atmósfera gaseosa primitiva de la tierra tomó lugar, en un recipiente experimental, cantidades enormes de residuos de alquitrán hubieran resultado, de esta manera la "tierra primitiva" habría tenido cantidades enormes de alquitrán de nitrogeno producido no biologicamente que se hubiera incorporado en los sedimentos de el Precambrico primitivo. No se conoce este alquitrán producido no-biologicamente en el registrogeologico. De esta manera también, debemos concluir que no tenemos ninguna evidencia de que el océano de "sopa diluida de pollo" existió alguna vez. Solamente un deseo de la imaginación es lo unico que le permite sobrevivir. Como alguien ha propuesto " El registro de la evolución biológica se manifiesta en la química de los organismos vivos....probablemente provee la evidencia más convincente para un período de evolución química temprana en la historia de la tierra." Esto es claramente redundancia.
Muchos autores apoyan la ausencia de "una sopa caliente diluida" A. G. Cairns-Smith (1982), W. Day (1984), H. D. Pflug (1984), C. R. Woese (1979), Hulett (1969), Shapiro (1986), M. Delbruck (1986), muchos de los cuales creen de alguna manera en un origen de vida pre-biologico, concluyen que no hay evidencia de que el proceso haya ocurrido. A pesar de esto, un número igual de autores consideran el escenario del origen de la vida tan bién establecido que no necesita justificación!. Sagan y M. J. Newman han ido más lejos al declarar que " la ausencia de la evidencia es la evidencia de la ausencia." Para nosotros que creeos que la vida no pudo haberse originado de una "sopa caliente diluida de pollo" que nunca existió, la declaración religiosa irracional de Sagan ha añadido más fortaleza a nuestra convicción de que él esta equivocado!
Hasta aquí hemos lidiado con el problema de pequeñas moleculas. Concluimos que la tierra no tuvo una atmósfera reductora y que aunque la hubiera tenido, no hay posibilidades de que ella hubiera dado origen a un océano lleno de pequeñas moleculas, requerido por los que proponen la evolución prebiotica para producir la primera célula. Ahora tenemos un camin largo que recorrer y yo todavía no he terminado. Asi mismo vamos a admitir la existencia de un oceano lleno de pequeñas moleculas y ver que podemos hacer con este.
El Surgimiento de Biopolímeros Ricos en Información:
Dado un oceano lleno de moleculas del tipo mas probablemente producido por una tierra pre-biologica, por los procesos sugeridos por los cientificos, debemos ahora considerar la pregunta de la polimerización. La pregunta de la polimerización es una espada de dos filos: debemos primeramente demostrar que la sintesis de macromoléculas es posible bajo condiciones prebiológicas,después debemos elaborar una manera lógica de generar macromoléculas ricas en información, necesarias para el desarrollo de una pre-célula.. Vamos a considerar las dos cosas separadamente.
La sintesis de proteinas y acidos nucleicos a partir de pequeñas moléculas precursoras, representa uno de los desafíos más difíciles del modelo de la evolución prebiológica. Todas las propociciones existentes presentan muchos problemas. Polimerization es una reacción en la cual agua en un producto. De esta manera va a ser entonces favorecida solo por la ausencia de agua. La presencia de precursores en un océano de agua favorece la despolimerización de cualquier molecula que puedan ser formadas. Experimentos cuidadosos hechos en una solución aquosa con altas concentraciones de aminoacidos demuestran la imposibildad de la polimerización significantiva en este ambiente. Un analisis termodinámico de una mezcla de proteinas y aminoácidos en un océano conteniendo una solución de 1 mol de cada aminoácido (esta concentración 100,000,000 veces más grande a la que se supone que existió en el océano pre-biótico) indica que la concentración de proteinas que contienen apenas 100 enlases peptidicos (101 aminoácids) en equilibrio seria de10-338 molar. Solo para hacer este número fácil de comprender, nuestro universo debe tener un volumen aproximado de 1085 litros. A 10-338 molar, necesitariamos un océano con un volumen igual a 10229 universos (100, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000, 000) solo para encontrar una sola molécula de qualquier proteina con 100 enlaces peptídicos. Por lo tanto debemos procurar de encontrar otro mecanismo para producir polímeros. Este no puede pasar en el océano.
Sidney Fox, un químico de aminoácidos, y uno de mis profesores, reconoció el problema y comenzo a elaborar una alternativa. Ya que el agua no favorece la formación de enlaces peptídicos, la ausencia del agua favorece la reacción. Fox trato de fundir aminoácidos puros y cristalinos a fin de promover la formación de enlaces peptídicos, retirando el agua de la mezcla. El descubrió para su consternación que la mayoría de los aminoácidos se rompian antes de que se fundieran, produciendo productos de alquitrán. Después de muchos intentos el descubrió que dos de los veinte aminoácidos, ácido aspártico y ácido glutamico, se fundian a más o menos 200oC. El también descubrió que si disolvieramos los otros aminoácidos en ácido aspártico y ácido glutamico ya fundidos podriamos producir una fundición conteniendo 50% de los 18 aminoácidos restantes. No fué entonces una sorpresa que el líquido de color amber, después de ser cocinado por unas pocas horas, contenía polímeros de aminoácidos que presentaban algunas propiedades de las proteinas. Posteriormente, Fox llamó estos productos proteniodes. El material polimerizado puede ser vaciado en una solución acuosa, resultando en la formación de spherules de material semejante a proteína, que Fox consideró semejante a celulas. Fox ha reclamado casi todas las propiedades imaginables para su producto, incluyendo que él había alcanzado la transición de la macromolécula para la célula. El fué más lejos al punto de demostrar que un pedazo de roca de lava pudiera substituir un tubo de ensayo en la síntesis de proteínas y afirmó que el proceso ocurrio en la tierra primitiva en los alrededores de los volcanes. Sin embargo sus críticos y sus propios alumnos desnudaron su credibilidad. Observe los siguientes problemas:
1) Proteinoides no son proteinas; ellos contienen muchos enlaces que no son peptídicos y enlaces cruzadosque no son naturales.
2) Los enlaces petídicos son del tipo beta, mientras que todos los enlaces peptídicos biológicos son del tipo alpha.
3) Los materiales con los que él inició en experimento fueron aminoácidos purificados, que no tenian semejanza con los matreriales disueltos en la "sopa organica". Si alguien tuviera que hacer el experiemnto con la "sopa pre-biotica¨, el único producto sería alquitran.
4) El porcentage de 50% ácido aspártico y ácido glutamico necesario para estos experimentos no tienen semejanza a el porcentaje muchisimo más superior de glicina y alanina encontrados en los experimentos de sintesis de la tierra primitiva.
5) No hay evidencia del contenido de información de las moleculas.
Todas las alegaciones expuestas por Fox fallaron en pasar las pruebas cuando fueron examinadas cuidadosamente. De la misma manera que sus resultados parecias prometer mucho en la teoría, la realidad fué catastrofica para las esperanzas de los paleobiogequímicos.
Un número de otros enfoques han sido propuestos. La más optimistas de estas es el uso de arcillas. Arcillas son muy finas, y muy bien organizadas de silicatos de aluminio complejos asociados con muchos otros cationes. En este ambiente, los grupos de aminoácidos tienden a ordenarse, y polimeros de dozenas de aminoácidos han sido producidos. Mientras que estos estudios han generado un interes de partes de los evolucionistas prebióticos, su relevancia ha sido sofocada por diversos factores.
1) A pesar de que los animoácidos estan ligados por enlaces peptídicos, el producto contiene información que no tiene sentido.
2) Las arcillas muestran preferencia por aminoácidos básicos.
3) Deben ser usados aminoácidos puros y activados, ligados a adenine, para que ocurra la polimerización. Aminoácidos adenilados no es el material flotante más comun en el océano pre-biotico.
4) No ocurre la polimerización de aminoácidos si aminoácidos libres son utilizados.
5) Los polimeros resultantes son en su mayoría tridimencionales y no lineares, como deben ser los biopolímeros.
Por lo menos un científico optimista (Cairns-Smith, 1982) cree que las própias partículas de arcilla fueron la sustancia que formaron al primer organismo! En la realidad, lo mejor que se puede esperar de tal escenario es un polímero racémico que contiene aminoácidos proteinicos y no proteicos, que no tenían la menor relevancia para los sistemas vivos.
Recientemente fué abierto un capítulo final con el descubrimiento de las moléculas de ARN autocatalíticas. Estas fueron recibidas con mucha alegria por los evolucionistas prebióticos porque daban esperanza de disminuir la necesidad de fabricar proteinas en la primera célula. Las mismas fueron llamadas "ribozimas" y probaron ser incapaces de responder a la situación debido a dos fatores: ellas estan muy limitadas porque no son capaces de producir los precursores de ARN por cualquier mecanismo prebiótico considerado hasta ahora es un problema tan grande el que las ribozimas pretenden resolver:
1) Mientras que una ribosa puede ser producida bajo las condiciones pre-bioticas simuladas a través de la reacción de formosa, esta es un azucar raro en los polímeros de formaldeído (mecanismo pre-biotico que se acredita dar origen a los azucares). A demás de la prersencia de sustancias de nitrogeno dichos aminoácidos en la mezcla de reacción podrian prevenir la sintesis de azucares (Shapiro, 1988).
2) Cuando una ribosa es producida y condensada con una base nucleotica, tenemos una mexcla de isomeros ópticos, y por lo tanto solo uno es relevante a los estudios pre-bioticos.
3) La polimerización de los nucleotidos es inhibida por la incorporación de tal enantiomorfo.
4) Mientras que solo los polimeros 3'-5' ocurren en los sistemas biologicos, polimeros 5'-5' y 2'-5' son favorecidos en las reacciones sinteticas de tipo prebiológico (Joyce and Orgel, 1993).
5) Ninguna de las 5 bases presentes en DNA/RNA son producidas durante la oligomerización HCN en soluciones diluidas (el mecanismo pre-biotico que se cree que dió origen a las bases nucleotídicas). Muchas otras bases no codificadoras competirian durante la polimerización en mayores concentraciones de HCN.
Además de los problemas de sintesis de los precursores y de las reacciones de polimerización, todo el bosquejo depende en la abilidad para sintetizar una molécula de RNA la cual es capaz de hacer una copia de si misma, una hazaña que hasta ahora ha eludido esfuerzos extremados. La molécula debe tambien realizar alguna funcion vital para iniciar la fuerza de la vida. Hasta ahora toda esta conversación de un " Mundo de RNA" permanece en nuestro deseos mejor categorizada como ficción. El punto más desvastador de este esquema es. sin embargo, que no ofrece pistas de como llegar desde este bosquejo de el mecanismo de las proteinas de ADN-ARN de todas las células vivas. El hecho de que algunos científicos deciden exibir tal entusiasmo por este esquema, revela que poca fé tienen en los otros escenarios del origen de la vida, incluyendo los que fueron discutidos anteriormente.
Interferancia del Investigador, Proximidad y los Estereoisomeros:
En mi intento para establecer credibilidad en varios modelos para el origen de la vida, yo he descuidado ciertas consideraciones de gran importancia, a fin de permitir que los experimentos fuesen analizados para sus propios méritos. Más ahora no podemos ignorar estas consideraciones.
En todos los estudios experimentales sobre el estudio de la vida, la presencia del investigador hace una contribución muy significativa a las concluciones y condiciones del mismo experimento. Cuando el investigador se propone a lograr un objetivo ( sintesis de los precursores o la polimerización de los precursores) el naturalmente se propone a definir un sistema que tenga alguna posibilidad de alcanzar la meta deseada. De esta manera se escojen las condiciones en las cuales los materiales sean apropiados para una tierra prebiotica, dandole a los resultados un aire de credibilidad. Las condiciones restantes son cuidadosamente manipuladas para alcanzar el objetivo deseado. De este modo, se deja al lector con la impresión de muchas cosas que no tenian probabilidad de ocurrir, habrian sido posibles en la tierra pré-biotica. Por ejemplo, cuando Fox realizó sus experimentos para producir proteinoides a partir de aminoácidos, usando roca de lava en lugar de tubos de ensayo, el dió la impresión de que este era un modelo plausible para la tierra pre-biotica. Lo que el estaba tratando de evitar enfatizar era que el estaba realizando la reacción en la lava caliente, usando una mezcla de aminoácidos cristalinos purificados producidos por un organismo biótico (grano de soya), y purificados por otro organismo biótico (hombre). El también controló cuidadosamente la temperatura, el tiempo de contacto con el agua. Dejo por su cuenta determinar cual sería el resultado de tal experimento que fuese llevado a cabo en una roca de lava caliente con una "sopa prébiotica" condensada.
Podemos hacer las mismas críticas de todos los estudios mencionados hasta aquí, desde el estudio clásico de Miller usando mezclas de gases purificados en un recipiente cerrado de vidrio, a los estudios con arcillas usando mezclas purificadas de aminoácidos adenilados. Muchos de estos fueron planeados para obtener los resultados deseados y no las condiciones que los mismos investigadores le assignaban a la tierra pré-biotica. Por eso, los resultados son usados para reenforzar la validez de la tierra abiótica que ellos nunca probaron. Los mismos que procuran trabajar con las condiciones abióticas no pueden impedir la influencia del investigador. Después de una revición cuidadosa del escenario de la investigación abiogénica, J. Brooks y G. Shaw (1973) concluyeron:
"Estos experimentos.... reivindacan la síntesis abiotica para lo que de hecho fué producido y designado por un hombre muy inteligente y bastante biótico".
Tal franqueza es agradable, honesta y bastande atrazada.
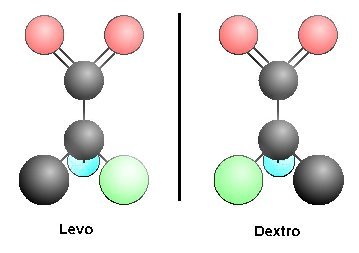
Otro problema igualmente serio es el de los enantiomeros o estereoisomeros. Este problema, es tal vez mayor que el anterior, porque frustra los esfuerzos de todos los investigadores pre-bioticos en alcanzar los resultados esencialmente significativos. Todos los compuestos de carbono que poseen cuatro grupos diferentes enlazados crean un centro de simetria. Esto permite que exista una molécula hermana formada por los mismos grupos con una configuración de imagen de espejo. Los dos compuestos formados presentan propiedades químicas identicas y generalmente pueden ser separados de su imagen hermana gemela optica usando filtros de sistemas biológicos.
|
||
Estos esteroisomeros presentan una dificultad para los investigadores de el origen de la vida. En los sitemas vivos, solo uno de los dos esteroisomeros es utilizado. Para los aminoácidos la forma utilizada es siempre la forma 1, para los azúcares la forma es d. Por eso , cuando las moléculas son sintetizadas en el laboratorio en las condiciones que se creen que habrian existidos en la tierra primitiva, ambos isomeros son formados en cantidades iguales. ¿Como entonces se puede explicar la predilección de escojer uno solo de los dos isomeros (en este caso los aminoacidos o isomero 1 para todos los 20 aminoácidos) cuando las propiedades son identicas para todos? Ya se han hecho grandes esfuerzos para intentar superar este problema, pero sin ningun exito. Se esperaba que tal vez las arcillas pudiesen discriminar los esteroisomeros, pero esto no sucedio. Esta dificultad no puede ser descartada. A menos que se encuentre una explicación para escojer los stereoisomeros , la unica solución para el problema es la que yo creo que es la correcta: la existencia de un diseño.
Otro desafio complicado para todos los esquemas que explican el surgimiento de polímeros informativos, tanto proteínas como ácidos nucleícos, es la presencia de reactivos competidores. Gould et al. (1981) sugerió que deben existir diez veces más aminoácidos no proteinicos de los que estan envueltos en la formación de proteinasn (20). Como se tiene poco o ningun control de las reacciones que ocurrieron, la presencia de especies competidoras , sean los aminoácidos no proteinicos o bases no codificadoras, llevará a la ocurrencia de darnos un desenvolvimiento no sistematico de moleculas informativas. Por lo tanto, si somos exitosos por un tiempo en el uso de bases y aminoácidos correctos, las concentraciones relativas de las especies competidoras aumentaran, empeorando el problema con el correr del tiempo. Cualquier calculo de probabilidad de originar alguna cosa que sera subestimado, debido a la incapacidad de cuantificar e incluir las contribuciones de estos elementos.
Un tercer desafio yo lo llamo "el problema de proximidad". Si un cierto número de moleculas son necesarias para producir una célula viva, producir una molécula en el Océano Indico y otra en el Océano Atlantico, no serviria de nada. El sistema de moléculas que formará una célula viva debe estar en el mismo lugar al mismo tiempo. Debido a esto, los sitemas asociados con particulas de arcilla fueron muy favorecidos en la comunidad abiogenica. Pero aunque las particulas de arcilla son catalizadoras, falta demostrar que todos los componentes necesarios para una célula podrian ser producidos espontaneamente en un mismo lugar. Naturalmente, hasta que demostremos cuales son los requerimientos mínimos que constituyen una célula viva, aquellos que quieran pueden continuar llamando las particulas de arcilla vivas. Yo tengo patrones superiores, debido a que los primeros modelos de células vivas parecen ser tan complejos como las formas modernas.
¿Puede la Vida Originarse de una "Sopa Orgánica"?
¿Que es la vida? Esta es una pregunta importante. Si existe en el mundo actual una continuación de parte de lo no vivo para lo vivo, entonces tal vez seria mucho dificil visualizar las cosas no vivas, ultrapasando la frontera. ¿Que constituye una célula viva? ¿Que criteria deben ser cumplidas?
1) Esta debe tener integridad- debe ser una entidad continua en si misma.
2) Esta debe ser limitada- debe tener un recipiente que sea parte de la entidad.
3) Esta debe ser capaz de reproducirse, su contenido y su recipiente.
4) Debe ser capaz de importar material y energia.
5) Debe ser capas de producir moléculas no derivadas de su ambiente.
Esta lista podria ser extendida infinitamente, más ahora nos vamos a fijar solo en estas. Analisemos la tercera propiedad, la de reproducción. ¿Cual es el nivel mínimo de información necesario para capacitar una célula para que se pueda reproducir? ¿De donde viene esa información? Varios enfoques pueden ser provistos para esta pregunta. Probablemente el enfoque más válido viene del estudió de el organismo menos complejo de todos los seres vivientes. Tal estudio proporciona estimados de varios centenares de millones de pedazos de información ( i.e. 100,000-1,000,000 nucleotidos). Otros insisten que un enfoque empirico reducionista sería más racional- lo que la célula necesita como mínimo para poder llevar a cabo la reproducción. Varios estimados dependen en gran parte del estado de optimismo del autor envuelto, pero excepto por los casos de extrema ingenuidad, los estimados giran en torno de 100 proteínas con funciones especificas de duplicación, transcripción y traducción. Fabricar proteinas de forma reproducible requiere información compleja, y esta información debería estar primeramente disponible en forma del ADN o ARN. Pero ya que el contenido de información de el ADN o ARN se aproxima a las proteínas producidas por ADN/ARN, los problemas son semejantes en ambos casos.
Debido a que el caso es más claro, consideremos primeramente los problamas asociados con la creación de una proteina, y particularmente, una única proteina o citocromo C. El citocromo C va a ser un ejemplo útil ya que esta distribuido en la naturaleza, y es la proteína más bien sequenciada. Debido a que esta presente en practicamente todos los organismos , ella tendria que estar entre las primeras proteinas celulares. El citocromo C consiste en una secuencia de más o menos 110 aminoácidos y los citocromos C de más de 100 organismos han sido secuenciados. Por lo tanto, para esta proteina , podemos tener una estimación razonable sofisticada de lo que sería necesario para producir una molecula funcional. En cada uno de los lugares diferentes de los 110 aminoácidos podemos determinar que sustituciones son permitidas a lo largo del espectro total de las proteinas secuenciadas. Por ejemplo, en la posición 93, el aminoácido presente puede ser Phe, Met, Ile, o Leu. Cada una de estas variaciones dan la proteina llamada citocromo C y es totalmente funcional, por lo tanto podemos decir que del uso de cualquier de estos cuatro aminoacidos en la posición 93 puede resultar una proteina totalmente funcional. Un calculo semejante para cada posición de los aminoácidos nos puede ofrecer una probabilidad minima, muy util, de obtener un cotocromo C de mutaciones al azar de aminoácidos. Calculos cuidadosos hechos por Hubert Yockey (1992) demuestran que una molecula de citocromo C funcional podria ser obtenida solamente en 2 x 1075 tentativas en caso de que todos los aminoácidos presentes estuviesen en cantidades equimolares y no hubiesen moleculas competidoras o esteroisomeros. Si uno acepta los estimados optimistas de Sagan de 1044 aminoácidos presentes en su sopa primitiva, y si pidieramos simultaneamente añadir un nuevo aminoácido a cada una de las 1044 cadenas en formación, por un segundo, prosiguiendo solamente hasta fallar, se necesitarian solamente 1023 años para tener una probabilidad de 95% de obtener una molecula fucional de citocromo C en este sistema. Esto es diez trillones veces más que la edad que es generalmente acceptada para el universo. El citocromo C, es una molécula muy liberal comparada con, digamos una proteina histona A3, que es tan invariante y que tiene solamente 3 aminoácidos diferentes entre la histona A3 del guisante de jardin y la del hombre. Para producir una única proteina histona correcta en este mismo sistema, serian necesarios casi 1060 añosa el nivel de 95% de probabilidad confiable, si solamente enlaces alfa fueren formados y 1-aminoácidos estuvieran presentes, y si no hubieran aminoácidos no-proteínicos competitivos en un sistema donde tales tentativas pudieran ser realizadas. En ámbas historias estamos suponiendo que tenemos este sistema, pero ya hemos visto que en la realidad esto no es así. En resumen, no es posible realizar la síntesis de proteínas o ácidos nucleicos conteniendo información relevante.
Hemos llegado a un callejon sin salida. Hasta este punto hemos analizado todo cuidadosamente, para tratar de encontrar una solución al dilema del origen de la vida. Aunque nosotros expandieramos los calculos de probabilidades con todas las substituciones funcionales posibles para una proteina que más conocemos, podemos ver que es practicamente imposible, aún en las condiciones más optimistas e irreales. ¿Como entonces podemos producir una célula viva? no pudimos conseguir producir una proteina funcional! Nos podemos detener aquí o seguir enterrando más los argumentos del origen abiogénico de la vida. Vamos a continuar!
Origen de las Células:
¿Que es necesario? La célula se define como una unidad viva capaz de auto-duplicarse, capaz de crecimiento, metabolismo y otras funciones asociadas con la vida. Vamos a enfocar el aspecto de la auto duplicación de la célula para que podamos determinar la posibilidad del surgimiento de una célula al azar. Si pudieramos visualizar los requisitos minimos, podemos entonces questionar si tal entidad seria capaz de auto-originarse. Los requisitos son formidables- primero necesitamos tener una información necesaria para la construcción celular, ya que sin información, la construcción celular de la propia vida es imposible. Todas las células vivas cuentan con información precisa acerca de su composición y división en la forma de ADN. El ADN es la representación molecular de la información codificada para el proceso de la estructura de la vida. Podemos argumentar sin base acerca de donde se origino esta información, pero la experiencia humana y los analisis cibernéticos nos confirman que la información provino de un "informador", creando así la necesidad de un dador de información. Un gran número de científicos muy respetados, incluyendo a el astrónómo Hoyle, el paleontólogo Patterson, o el cibermenicista Yockey y otros llegaron a conclusiones semejantes por razones muy distintas. Aun así persiste la creencia de que si hubiesen condiciones adecuadas y la cantidad de tiempo necesaria, cualquier cosa sería posible. Analizaremos entonces tal proposición para probar su validez.
Vamos a preguntar cuales son los requisitos mínimos para poder obtener una célula viva. Todas las células necesitan tener una membrana, en los casos más simples, de lípidos del tipo de triglicéridos o fosfoglicéridos asociados a proteinas especializadas, que estabilizan la membrana y aseguran su integridad estructural. Podemos observar la formación espontanea de dos capas de origen lípido en estructuras esféricas semejantes a las células. Así mismo podemos concluir que la presencia de fosfolípidos en el mar pre-biotico, aseguraría que el envase de la célula estaba presente. Pero el caso no es tan simple. Es extremadamente difícil de producir ácidos grasos, o componentes primarios de todas las membranas celulares, en las condiciones abiogénicas y en atmósferas reducidas. Aunque las mismas moléculas fuese producidas, cationes bivalentes tales como Mg++ y Ca++ se combinarian con los ácidos grasos haciendo que se precipitaran a el fondo del mar y fuesen incorporados a los sedimentos Precambricos. Así mismo, si hubiesen sido inicialmente formados, ellos no estarían disponibles para la formación de las membranas. Estas son moléculas complejas que ciertamente no serian nada comun bajo las condiciones primitivas de la tierra. La existencia de una membrana celular no es igualmente muy comun. Pero el problema va más alla, ya que las membranas fosfolípidas son impermeables a la mayoria de las moléculas que la célula requiere para crecer. Las membranas de las células modernas evitan este problema teniendo como componente integral proteinas muy sofisticadas , que admiten selectivamente las moléculas deseadas. Claro que no se concibe que tales proteínas estuviesen a disposición de la primera proto-célula. De la misma manera, la existencia de una membrana celular retardaria el desenvolvimiento de una proto-celula, porque sin una membrana no puede haber célula. Otra pregunta muy compleja. ¿Y Ahora?
Vamos a tratar otro enfoque, olvidense de la célula y de la membrana -- ¿Que sería necesario, pero no mínimo para producir una molécula de proteína? Podriamos inmaginar proteínas menores que las actuales, digamos que con 100 aminoácidos de largo, utilizando menos de los 20 aminoácidos protéicos, un sistema primitivo de polimerase, tal vez apenas 100 proteínas específicas en total o quizas solo 80. Vamos a suponer que los aminoácidos no proteícos pueden ser utilizados, y tambien los enantiomeros. Todas estas suposiones son ridículas. No tenemos los materiles para comenzar, ni los materiales correctos. No tenemos idea alguna de como podríamos producir un polimero de 100 aminoácidos bajo las condiciones pre-bioticas. No hay posibilidad alguna de que estas condiciones extremadamente desfavoralbes pudiese producir un sistema capaz de autoreplicarse. Pero ahora que estamos en este negocio, vamos a hacerlo aun peor. De las 80 proteínas que dijimos que eran necesarias, vamos a permitir que las primeras 60 tengan cualquier secuencia de aminoácidos. De las 20 proteínas restantes, la primera tiene un aminoácido específico, y los 99 restantes pueden ser de cualquier otro tipo. La segunda tiene 2 aminoácidos específicos y así sucesivamente, hasta que la vigésima tenga 20 aminoácidos específicos. Permitiremos que el océano tenga 6 Km de profundidad, cubriendo toda la tierra, y que la concentración de aminoácidos sea de 1 molar para cada tipo. Dividiremos el océano en parcelas de 1 litro y vamos a considerar la hazaña realizada cuando un litro produce todas las proteínas necesarias . Pernitiremos que las proteinas sean producidas a una velocidad de un millos de tentativas por segundo en un litro. Vamos a admitir la misma probabilidad para los ácidos nucleicos. Con todas estas suposiciones hechas a favor para producir nuestra célula primitiva extremadamente liberal, no conseguiremos alcanzar este objetivo, con un 50% de probabilidad, una vez en 10186 años.
Naturalmente esta cifra es incomprensible. Para tener una idea de cuan incomprensible es, voy a usar seguidamente una ilustración. Una ameba esta de un lado del universo y comienza a caminar para el extremo opuesto, distante de 100 trillones de años luz. Ella viaja a una velocidad de un metro por cada billón de años. y carga con sigo un átomo. Cuando ella llega a el otro lado, descarga el átomo y comienza a caminar de vuelta. En 10186 años, la ameba habrá transportado toda la masa del universo de un lado para el otro trillones trillones trillones trillones trillones trillones de veces. Esta es mi definición de algo imposible, y lo que hubiera resultado exitosamente, no seria una célula viva, ni una combinación prometedora. El origen espontaneo de la vida en una tierra pre-biotica es IMPOSIBLE!
Alternativas para la Abiogénesis:
¿Cuales son las alternativas? Muchas han sido propuestas:
1) Origen en otro planeta. ¿Como ayuda esto? Ya hemos investigados las mejores condiciones posibles y descubrimos que no son productivas. Jugar con el problema de un lado al otro es admitir el fracaso.
2) Predestinación Bioquímica: Un esfuerzo de atribuir las propiedades de los sitemas vivos a las moléculas de las cuales estan formados. Un libro popular de 1969 con este título sugería este escenario. Esta linea ha sido abrazada por los teóricos y sociobiologistas. Definitivamente, la vialidad de un modelo no puede depender de un contenido atractivo, pero de cuan veráz es. No existe evidencia de que los precursores biologicos se han energetizados para producir células vivas. Uno de los autores del libro, Dean Kenyon, ahora es un creacionista.
3) La creación por un poder inteligente que esta afuera de nuestra esfera de investigación. Esta posibilidad es mejor investigada cuando consideramos las alternativas que existen. Ya hemos hecho eso. Ciertamente alguien que esta investigando por la verdad, no puede excluir arbitrariamente esta posibilidad.
A la luz de estas alternativas el concepto de la creación se torna extremadamente atractivo, no como una alternativa, pero como la única alternativa razonable. Si alguien no quiere admitir la posibilidad de una inteligencia superior excluiría esta consideración. Esta consideración por lo menos tiene sentido a la luz de muchas observaciones que consideramos, porque explica la fuente de información , la sincronicidad de los esteroisomeros, el arreglo no casual del código genñetico y las respuestas a los misterios más insondables que vinieron primero, las proteínas del código genético. Los cientificos deben darle la bienvenidad a una solución que trae entendimiento y orden a el caos. Los científicos deben ser los primeros en darle la bienvenida a su Creador!
Referencias
Abelson, P. H. (1966). Chemical events on the primitive Earth. Proceedings of the National Academy of Sciences USA 55 1365-72.
Brooks, J. & Shaw, G. (1973). Origin and Development of Living Systems. London; New York: Academic Press.
Cairns-Smith, A. G.. (1982). Genetic Takeover and the Mineral Origin of Life. Cambridge; London; New York: Cambridge University Press.
Day, W. (1984). Genesis on Planet Earth. New Haven; London: Yale University Press.
Delbruck, M. (1986). Mind from Matter? An Essay on Evolutionary Epistemology, eds G. S. Stent, E. P. Fischer, S. W. Golomb, D. Presti & H. Seiler. Palo Alto; Oxford; London: Blackwell.
Gould, S. J., Luria, S. E., and Singer, S. (1981). A View of Life. Menlo Park, California: Benjamin/Cummings.
Hulett, H. R. (1969). Limitations on prebiological synthesis. Journal of Theoretical Biology 24, 56-72.
Hull, H. E. (1960). Thermodynamics and kinetics of spontaneous generation. Nature 186, 693- 4.
Joyce, G. F. And L. E. Orgel (1993). Prospects for understanding the origin of the RNA World. In The RNA World, ed. R. F. Gesteland and J. F. Atkins. New York: Cold Spring Harbor Press.
Lazcano A, and S. L. Miller. 1996. The Origin and Early evolution of life: Prebiotic chemistry, the pre-RNA world, and time. Cell 85:793-798.
Pflug, H. D. (1984). Early earth geological record and the origin of life. Naturwisschenschaften 71, 63-8.
Schidlowski, M. (1976). Archean atmosphere and evolution of the terrestrial oxygen budget. In The Early History of the Earth, ed. B. F. Windley. London; New York: John Wiley & Sons.
Schopf, J. W. (1972). Precambrian Paleobiology. In Exobiology, ed. C. Ponnamperuma. Amsterdam; London: North-Holland.
Shapiro, R. (1986). Doubt and uncertainty; Bubbles, ripples and mud. In Origins, A Skeptic’s Guide to the Creation of Life on Earth, Chapter 8, pp. 190-224. New York: Summit Books.
Shapiro, R. (1986). Prebiotic ribose synthesis: a critical analysis. Origins of Life 18, 71-85.
Sillen, L. G. (1965). Oxidation state of Earth’s ocean and atmosphere, Arkiv for Kemi 24, 431- 56.
Walker, J. C. G. (1976). Implications for atmospheric evolution of the inhomogeneous accretion model of the origin of the Earth. In The early History of the Earth, ed. B. F. Windley. London; New York: John Wiley & Sons.
Walker, J. C. G. (1977). Evolution of the atmosphere. New York: Macmillan.
Woese, C. R. (1979). A proposal concerning the origin of life on the planet Earth. Journal of Molecular Biology 13, 95-101.
Yockey, H. P. (1992). Information theory and molecular biology. Cambridge; New York; Oakleigh: Cambridge University Press.
Yuasa, S., Flory, D., Basile & Oro, J. (1984). Abiotic synthesis of purines and other heterocyclic compounds by the action of electrical discharges. Journal of Molecular Biology 21, 76-80.