![]() Hominid skulls
Hominid skulls
APES, APEMEN AND MEN
.A Creationist perspective
Ó
2004 Lee A. Spencer, Ph.D.In the modern world, there are two competing theories for man¢ s origin, (Photo of ark) naturalism/evolution and theism/creationism-diluvialism. The purpose of this particular paper is not to refute the concept that man and modern apes evolved from a common ancestor. In principle, anyone reasonably acquainted with the marvels and usefulness of the human mind ought to be looking to an explanation other than evolution on that ground alone. We do, however, wish to make it abundantly clear that we do not believe the hominid fossil record to be the result of evolution. Even within the evolutionary community, there are no hypotheses of descent that are not falsified by the appearance of some characters out of sequence. This will be abundantly documented in a separate forthcoming paper. This paper is being presented in an effort to lay out the evidence revealed by the fossils, that we may begin to consider explanations with a full deck.
Historically, virtually all modern creationist writers, if they address the subject at all, state that the fossil ape-men are either degenerate, fully modern humans or simply extinct apes; there are no transitional forms. Lubenow (1989) developed his book on the basis of these premises. He defines skull 1470 and all of the other skulls with human-like morphology as fully modern man. He defines the Australopithecines and Homo habilis as simply extinct apes. Thus he effectively defines away the problem of transitional features. It is true as a generalization, that transitional morphologies between major groups of biological organisms are lacking. However, is this true for the hominids? How would we recognize transitions if they were present? And what would they signify? The focus of this article is to test whether morphological transitions are preserved in the fossil record of hominids, to consider how a transition would be identified, and to attempt to reach some conclusion regarding transitional forms in the hominid fossil record. Do any of the fossil hominid species show evidence of being diseased or degenerate modern humans, as Lubenow and others assert, or are there transitional morphologies that need to be explained?
There are three families of hominids represented by living forms today. These family groups are characterized on the basis of mode of locomotion. Thus man, the only living primate that habitually walks upright, and is placed in his own family, the Hominidae. The knuckle-walking apes (gorilla, chimpanzee, and orangutan) are in the family Pongidae. The gibbons are brachiators (swinging arm movements) and are placed in the family Hylobatidae.
We will begin by examining the cranial specimens from members of the families Hominidae and Pongidae. Then we will consider the post-cranial data.

![]() Hominid skulls
Hominid skulls
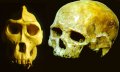
Living anthropoids exhibit distinctly different skull morphologies. In the figure to the right is pictured an adult male gorilla skull. It can be characterized as having a prominent saggital crest (ridge along the top of the skull), a large bony orbit (eye socket), and a prominent brow ridge. It also has a large flat facial area below the orbits and a protruding muzzle (prognathism). Compared to the gorilla skull, the human skull lacks a saggital crest, has gracile orbits (thinner and smaller), and a short face, with no prognathism. There is no confusion about what is human and what is ape; there are no morphologic intermediates.
However, if the fossil hominids are included, a different picture emerges. A gradation of morphological characters emerges ranging from those features that are mostly chimp-like to those that equal modern humans. These character states may be seen in endocranial volumes, various cranial features, facial structure, placement of the foramen magnum, and construction of the face and mouth.
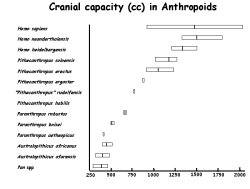
Fully modern man can be characterized as having a brain capacity that averages about 1700 cc. Conversely; the brain capacity of the pongids (gorilla, chimpanzee, orangutan) averages about 400 cc. There is no overlap between the smallest human brain capacity and the largest simian brain capacity. However, as the figure to left illustrates, if we plot values including the fossil hominids, a continuum of endocranial volumes between the chimpanzee and modern man results. There is a gradation in endocranial capacities in the hominids, extending from the great apes to man. One consideration that should not be overlooked in assessing endocranial measurements is the significant direct correlation between skeletal size and endocranial capacity. The range of brain sizes in the modern population is a predictor of the range in body sizes in the same population. But the mean values themselves (dark vertical bars) are still gradational.
![]() Anterior View
Anterior View
 .
.
1 Chimpanzee 2 A. africanus 3 H. habilis 4 KNM-ER 1470 5 Java Man 6 Peking Man 7 H. saldensis 8 H. saldensis 9 Broken Hill 10 Neanderthal 11 Modern Man
A number of features can be compared from the anterior aspect of the skulls (figures are hyperlinked to larger photos).In the group of skulls illustrated above, note that several chimpanzee (1) skull features are maintained up through and including the Neanderthal (10). These features include a pear-shaped cranium when viewed from above, as opposed to the distinctly oval aspect of the modern human (11), resulting in part from a pronounced narrowing in the temporal region; a prominent brow ridge, seen in all of the above except for modern man, and a protruding muzzle, most evident in those skulls which have faces preserved.
![]() Dorsal View
Dorsal View

1 Chimpanzee 2 A. africanus 3 H. habilis 4 KNM-ER 1470 5 Peking Man 6 OH 9 7 H. saldensis 8 H. saldensis 9 Broken Hill 10 Neanderthal 11 SKUL 12 Modern Man
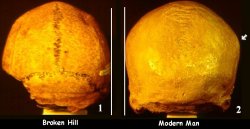
The increase in endocranial volume can also be seen in the dorsal view. Once again, several features prominent on the Chimpanzee skull (1) are also found on the other hominid skulls through Neanderthal (10). Notice that in all of the skulls beginning with the chimpanzee and extending through H. neanderthalensis, the widest part of the skull is at the base. This is in marked contrast to the skull of H. sapiens, in which the widest part of the skull is in the parietal region (see photo to left comparing Broken Hill skull with that of modern man). This prominent feature is called the parietal eminence. The parietal eminence is not found in any hominid other than fully modern man. Another feature more obvious in the skulls themselves than it is in this photo is the presence of the attachment scars for the neck musculature. These scars are also prominent features on all skulls from chimp to H. neanderthalensis where the muscle attachment often extends quite high on the back of the skull (see below).
![]() Lateral View
Lateral View
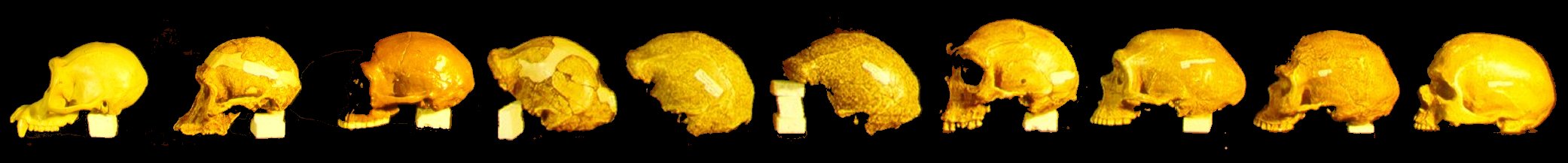
1 Chimpanzee 2 A. africanus 3 Peking Man 4 OH 9 5 H. saldensis 6 H. saldensis 7 Broken Hill 8 Neanderthal 9 SKUL 10 Modern Man

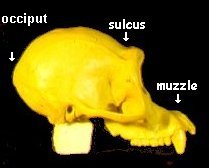
Beginning with the Chimpanzee (1, and to left), notice the protruding muzzle, the sulcus (groove) behind the orbital ridge, and the pointed termination of the occiput at the mid level of the dorsal surface of the skull. This feature is expressed in the occipital bun in the Neanderthal (8, and to right), but is completely missing in the modern skull. Notice also the muscle attachment scars are responsible for the dihedral aspect of the dorsal surface. All of these features can be seen in the Neanderthal skull. Notice that the Australopithecines and homo-erectines are intermediate in expression of these features, and that they are absent in the modern skull.
![]() Frontal View
Frontal View

1. Chimpanzee 2 A. africanus 3 Peking Man 4 Steinheim Man 5 Neanderthal 6 Fully modern human
From the frontal view, all of the specimens other than fully modern human (6) share the same basic facial structures of large orbits, with heavy bony eye rings, a prominent zygomatic arch, and nares nearly the size of the orbits. Fully modern man has proportionately smaller orbits, and a much more gracile facial structure that lacks the bony eye rings and has proportionally smaller nares.
![]() Posterior View
Posterior View
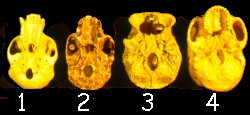
From the posterior view, two key characteristics relating to morphologic transitions are apparent. The chimpanzee (1) and indeed, all of the pongids (bonobo, chimp, gorilla, and orangutan) are knuckle-walkers. The orang makes a fist with the first phalanx bearing the weight. In all of the other pongids, it is the second phalanx that bears the weight. Because they are all knuckle-walkers, the opening for the spinal cord (foramen magnum) must be located farther back on the skull ( posteriorly) so that the head can be rotated to lift the eyes to the direction of travel. In fully modern humans, the skull rests directly over the axis of the body, so the body bears the weight of the head. The eyes already point in the direction of travel. Therefore, the foramen magnum is nearly in the center of the skull. Notice the placement of the foramen magnum in these four skulls. The australopithecine and Homo erectus are distinctly intermediate with respect to the position of the foramen magnum, between the chimpanzee and fully modern human.
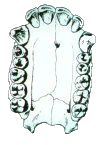
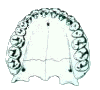
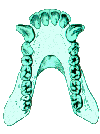
The other key characteristic seen in this view is the shape of the palate. The pongids have a rectangular shaped palate with very large canine teeth (left picture). In order to be able to close their mouth, there must be a gap (diastema) for the opposing tooth between the incisors and the canine. The fully modern human palate is parabolic in shape, rather than rectangular (right picture). The canines are relatively smaller so that the teeth can occlude without a gap for their placement.
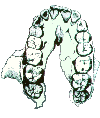

The palates of the australopithecines (right picture), and to a lesser extent the homo-erectines (H. erectus s.l., left picture) are intermediate in shape between the chimpanzee and fully modern humans. The tooth row in the fossil hominids tends to remain fairly straight, but tapers toward the incisors. The incisors have a general parabolic shape.

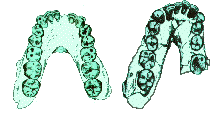
The lower jaw (mandible) must have the same general shape as the palate in order for the teeth to occlude. The rectangular shape of the pongid palate is represented in the mandible as well, including the diastema (left picture). Once again, the pongids have a rectangular shaped tooth row, whereas modern humans (right picture) have a parabolic mandible shape. The pongid mandible has a bony extension lingual to the incisors, called the ² simian shelf.² Fully modern humans lack this bony extension.
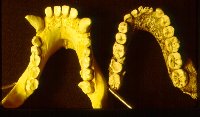
The mandibles of the australopithecines (right picture), and to a lesser extent the homo-erectines (H. erectus s.l., left picture, with gorilla for comparison) are also intermediate in shape between the chimpanzee and fully modern humans. The tooth row in the fossil hominids tends to remain fairly straight, but tapers toward the incisors. The incisors have a general parabolic shape. Notice that the homo-erectines have a simian shelf that is about half the length of that of the pongids. All of the fossil hominids, including Neanderthal have a simian shelf, though it may be reduced in size. Fully modern humans have no simian shelf. To have a simian shelf is an ape characteristic; to lack a simian shelf is a human characteristic.

The final difference in jaw shape can be seen in lateral aspect. Fully modern human mandibles exhibit a bony protuberance on the angle of the chin referred to as the mental eminence (picture to right). This feature is entirely lacking in the pongids (picture to left), and fossil hominids, including Neanderthal. To have a mental eminence is a human characteristic; to lack a mental eminence is an ape characteristic.
![]() Postcranial Anatomy
Postcranial Anatomy
![]() Digits
Digits
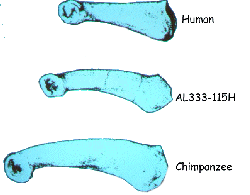
The figure to right illustrates proximal phalanges from a human, a chimpanzee and an Australopithecene. The curvature of the bone is considered to be an adaptation for grasping and swinging. The chimp bone is large with considerable curvature. The human is smaller and straight. The Australopithecene bone is intermediate in size and curvature.
![]() Pelvis
Pelvis
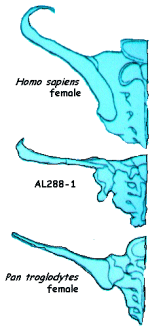
Three cross sections of the pelvic girdles of a modern human, an Australopithecene and a Chimpanzee are illustrated to the left. Notice the Australopithecene characteristics are more or less intermediate between those of the modern human and those of the chimpanzee. For most of the various parameters of the post-cranial skeleton (and hundreds of measurements are made), the human and chimp are well separated. The various hominid species are sometimes intermediate, sometimes fall well within the range of the human measurements, and sometimes fall with the apes. For example, for the metacarpels, the australopithecenes fall easily within the length measurements for modern man, and outside the range for the chimp. The same is true for the A-P width of the metacarpel II facet. But the styloid projection and waisting measurements find the ausralopithecenes well within the range of the chimps, or for some specimens, intermediate between human and chimp.
Thus, postcranial characters show the same combination of ape-like, human-like and intermediate character states for the pelvis, humo-femoral index, curvature and length of the phalanges (hand and foot), and wrist bones. We can conclude that the fossil hominids are cumulatively intermediate between modern apes and modern humans in the postcranial characters as well.
![]() Summary
Summary
The fossil hominid crania show a gradation of morphologies between those characteristics seen in the apes but not in humans, to those characteristics seen in humans but not in apes. There is also a gradation in endocranial volume. Similarly , the postcranial skeletons of the hominids exhibit many characteristics that are intermediate, as well as a mixture of other characteristics that are either ape-like or human-like. Therefore, it is difficult to maintain the position that there are no morphological intermediates between apes and man. So what explanation for the existence of fossil intermediates can we posit other than evolution from a common ancestor, an explanation that is falsified by the order of appearance of various features in various lineages? With the introduction of the phylogenetic systematic methodology, there are no unfalsified hypotheses of human evolutionary lineages. An explanation that is consistent with the data and with the Biblical account of origins is that man has been engaged in exchanging genetic material with the pongids, either in nature or in the laboratory. The variety of forms can be attributed to some form of reticulation wherein the original creatures outcrossed to both H. sapiens and pongids, as well as interbreeding within their own group. Michael Day, in a booklet in the Oxford/Carolina Reader series entitled "The Fossil History of Man" asserts that the skull from the cave of Arago "...shows a mixture of both erectus and sapiens features. It may be regarded as an example of mosaic evolution that may be an erectus/sapiens intergrade." That such is possible is doubted by few knowledgeable anthropologists. For example, Wong states, with respect to modern humans interbreeding with Neanderthals:
"Radovcic draws my attention to a grayish green band, the so-called G3 level that contained some of the Neanderthal fossils he himself unearthed and fishes a cast of one of the ancient bones out of his pocket. 'The Vindija hominids were modernized Neanderthals,' he says, showing me the partial lower jaw featureing the beginnings of a chin--one of the hallmarks of modern human morphology. And although other fossils from the site reveal typical Neanderthal traits such as the pronounced browridge, they are most delicate and modern in shape in the Vindija people than in earlier Neanderthals. Radovcic and others who have studied these remains believe this apparent shift toward the modern condition suggests interbreeding between Neanderthals and moderns--a case that is strengthened by early modern human fossil finds from central Europe that bear some Neanderthal-like features." Kate Wong, "Cave Inn," Scientific American, Dec. 1999, p. 34
That fossil hominids represent the results of hybridization between modern humans and apes is an hypothesis that should not be tested. And the moral implications of such an experiment are enough to keep most rational scientists from doing so.