SCIENCE AND FAITH: THE HOMINID FOSSIL RECORD
Lee A. Spencer, Ph.D.
Earth History Research Center
Southwestern Adventist University
Keene, TX 76059
Abstract— The fossil hominids are bipedal primates with a mixture of great ape (pongid), human, and pongid-human intermediate character states. There is a morphological continuum in skull and skeletal morphologies spanning those seen in the chimpanzee and anatomically modern man. Evolutionary/theistic evolutionary, progressive creation and “young-earth” creation interpretations of the data set are examined and tested. No interpretation remains unfalsified. Any interpretation held requires the exercise of faith. Science and faith not only are possible together, they are required.
Introduction
The cosmic questions of who we are and what our purpose in life is, if there is one, have been debated for as long as man has been in existence. So has the question of our origin. For millennia it has been commonly believed that a supernatural being or beings created man and this world. Over the last 150 years, this belief has been replaced by a naturalistic model in which man and this world came into being exclusively through naturally occurring processes. A conflict has ensued between those who believed in a supernatural being and those who do not. Few topics within the origins controversy engender more interest than that of man’s origin. Few topics better illustrate the intertwining of data and interpretation, science and faith, or cosmology and science.
Associated with the origins controversy, there is intense debate about what is or is not science, what constitutes religion and faith and how these factors combine to produce our cosmology. Because there are widely differing views on exactly what constitutes each of these terms, this paper uses the following definitions:
1) Data are directly observable characters and measurements. For the discussion of human origins the data set would include observations and measurements of skeletal remains, descriptions of sedimentary context, various chemical constituents, etc.
2) Interpretations are conclusions drawn from the data. For example, based upon microscopic grooves on the teeth (data), dietary modes may be inferred; based upon a measured ratio of 40K/40Ar, the number of calendar years required to produce that ratio may be hypothesized, etc. Some interpretations may appear to be self-evident. Conversely, there may be different and opposing interpretations of the same data set. Often in the scientific literature, data and interpretation are so intermingled that it is difficult to determine the actual data set, or only the interpretations are reported.
3) Science is the process of formulating testable (i.e. rejectable) hypotheses about the natural world.
4) Faith is the acceptance of statements, interpretations and/or hypotheses as true in spite of the lack of data or even the presence of contradictory data.
5) Cosmology is the belief system explaining the cause for the underlying harmony in the universe. Examples of cosmologies include naturalism where natural law is self-existent and is the sole cause for the underlying harmony of the universe, and variations of supernaturalism where there is a being above natural law. Evolution is the extension of naturalism that covers change through time. Theistic evolution, progressive creation, intelligent design, young earth creation, etc. are all variants of the supernaturalistic cosmology with differing levels or timings of intervention by the supernatural being. I know of no way to differentiate evolution and theistic evolution from observable data, so they will be treated together. Because cosmologies are belief systems, they are not directly testable and are hence outside the definition of science given above.
It is not the purpose of this paper to resolve the cosmological debate on human origins, but to show how our understanding of what man is depends completely… not on the data, but upon previously adopted views that form a previously accepted cosmology. Further, the exercise of faith is required for all cosmologies. We will first examine the human origin data set very briefly, then examine interpretations and hypotheses of the three most prevalent cosmologies: evolution/theistic evolution, progressive creation and neodiluvialism/young-earth creation.
A Summary of the Hominoid Data Set
(Details, Details, Details)
What Is A Hominid?
 Linnean
classification places all the apes with man in the superfamily Hominoidea. The
Hominoidea can be distinguished from all other primates in lacking a tail and
in having a wrist design that allows greater wrist flexibility and greater
dexterity of the hands and feet than other primates. The Hominoidea is divided
into three families, the Hylobatidae (gibbons), the Pongidae (orangutan,
gorilla, and chimpanzee), and the Hominidae (man). The three families are
distinguished from each other by different modes of locomotion and the
anatomical structures necessary for that type of locomotion.
Linnean
classification places all the apes with man in the superfamily Hominoidea. The
Hominoidea can be distinguished from all other primates in lacking a tail and
in having a wrist design that allows greater wrist flexibility and greater
dexterity of the hands and feet than other primates. The Hominoidea is divided
into three families, the Hylobatidae (gibbons), the Pongidae (orangutan,
gorilla, and chimpanzee), and the Hominidae (man). The three families are
distinguished from each other by different modes of locomotion and the
anatomical structures necessary for that type of locomotion.
 The
Hylobatidae (also known as the “lesser apes” or gibbons) are brachiators, which
means that they move by swinging arm movements with the body hanging below the
arms.
The
Hylobatidae (also known as the “lesser apes” or gibbons) are brachiators, which
means that they move by swinging arm movements with the body hanging below the
arms.
 The
Pongidae or “great apes” are forelimb knuckle-walkers. The orangutan makes a
fist and the first phalanx makes contact with the ground; the gorilla and
chimpanzee use the second phalanx to make ground contact. The term “pongid”
will be used throughout this paper to refer to this group.
The
Pongidae or “great apes” are forelimb knuckle-walkers. The orangutan makes a
fist and the first phalanx makes contact with the ground; the gorilla and
chimpanzee use the second phalanx to make ground contact. The term “pongid”
will be used throughout this paper to refer to this group.
The Hominidae (human beings) walk habitually upright and the hands do not normally touch the ground. Upright walking primates are known as “hominids” from the family name Hominidae and modern man, Homo sapiens, is the only living representative.
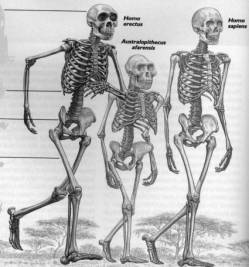 In
the fossil record, there are a number of extinct, upright-walking primate taxa
(hominids). The taxonomy of the fossil hominid specimens is hotly debated.
Depending upon the author, there are nine to eighteen species placed within two
to six genera. There is a general consensus, however, that all of the specimens
can be placed within one of six general categories: the australopithecines,
early “Homo”, the “erectines”, “archaic sapiens”, Neanderthals, and fully
modern man. The australopithecines and early “Homo” are restricted to the
African uppermost Miocene to Pliocene; the “erectines”, to the Old World
Pliocene and Pleistocene; “archaic
sapiens”, and Neanderthals to the Old World Pleistocene, and fully modern man
to the Pleistocene and Holocene worldwide.
In
the fossil record, there are a number of extinct, upright-walking primate taxa
(hominids). The taxonomy of the fossil hominid specimens is hotly debated.
Depending upon the author, there are nine to eighteen species placed within two
to six genera. There is a general consensus, however, that all of the specimens
can be placed within one of six general categories: the australopithecines,
early “Homo”, the “erectines”, “archaic sapiens”, Neanderthals, and fully
modern man. The australopithecines and early “Homo” are restricted to the
African uppermost Miocene to Pliocene; the “erectines”, to the Old World
Pliocene and Pleistocene; “archaic
sapiens”, and Neanderthals to the Old World Pleistocene, and fully modern man
to the Pleistocene and Holocene worldwide.
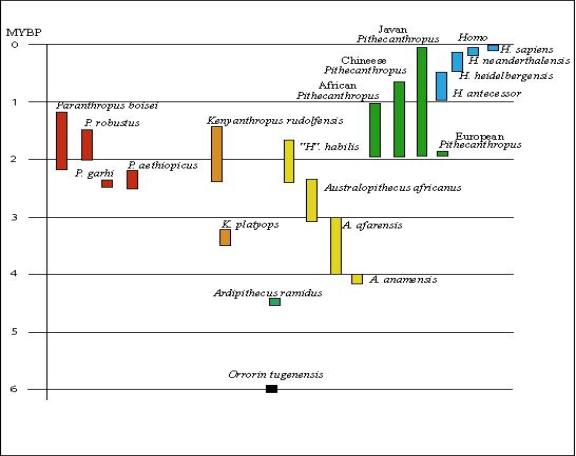
The classification of the hominid taxa used for this paper is modified from Tattersall (1993) and Wood and Collard (1999) and is as follows:
Family Hominidae
Insertae sedis Orrorin tugenensis
Subfamily Australopithecinae
Genus Ardipithecus
Species Ardipithecus ramidus
Genus Australopithecus
Species Australopithecus anamensis
Australopithecus
afarensis
Australopithecus
africanus
Australopithecus
bahrelghazali (?)
Genus undescribed
Species “Homo” habilis
Genus Paranthropus
Species Paranthropus aethiopicus
Paranthropus boisei
Paranthropus
robustus
Paranthropus garhi
Genus Kenyanthropus
Species Kenyanthropus platyops
Kenyanthropus rudolfensis
Subfamily Homininae
Genus Pithecanthropus
Species Pithecanthropus erectus
Pithecanthropus soloensis
Genus Homo
Species Homo heidelbergensis
Homo
neanderthalensis
Homo
sapiens
Data Set I: Hominoid Skull Characters
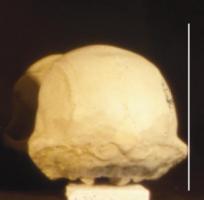
 Living
hominoids exhibit distinctly different skull morphologies. In the figure to the
left is pictured an adult male gorilla skull. It can be characterized as having
a prominent sagittal crest (ridge along the top of the skull), a large bony
orbit (eye socket), and a prominent brow ridge. It also has a large flat facial
area below the orbits and a protruding muzzle (prognathism) with large canine
teeth.
Living
hominoids exhibit distinctly different skull morphologies. In the figure to the
left is pictured an adult male gorilla skull. It can be characterized as having
a prominent sagittal crest (ridge along the top of the skull), a large bony
orbit (eye socket), and a prominent brow ridge. It also has a large flat facial
area below the orbits and a protruding muzzle (prognathism) with large canine
teeth.
 Compared
to the gorilla skull, the human skull lacks a saggital crest, has gracile
orbits (thinner and smaller), and a short face, with no prognathism. There is
no confusion about what is human and what is ape; there are no living
morphologic intermediates.
Compared
to the gorilla skull, the human skull lacks a saggital crest, has gracile
orbits (thinner and smaller), and a short face, with no prognathism. There is
no confusion about what is human and what is ape; there are no living
morphologic intermediates.
However, if the fossil hominids are included, a different picture emerges. A gradation of morphological characters is seen ranging from those features that are mostly gorilla and chimp-like to those that equal modern humans. Character states examined here include endocranial volume, various cranial features and facial structure, placement of the foramen magnum, and construction of the face, mouth and teeth.
Endocranial Volume. Fully modern man has a brain capacity that averages about 1700 cc. The brain capacity of the pongids (gorilla, chimpanzee, orangutan) averages about 400 cc. There is no overlap between the smallest human brain capacity and the largest simian brain capacity. However, if we plot values including the fossil hominids, there is a gradation in endocranial volumes.
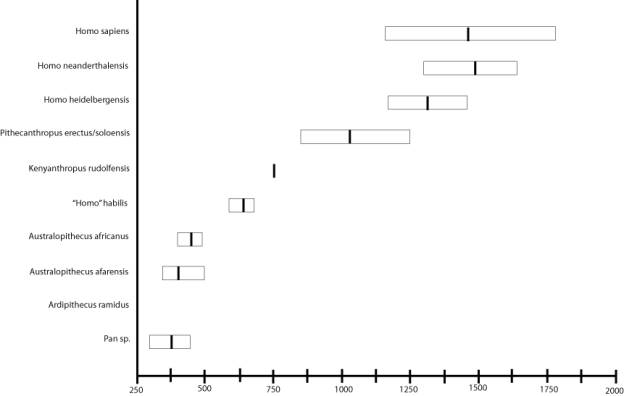

![]()
Cranial Morphology. Among the fossil hominids, we also see a gradation of cranial morphologies from an essentially pongid morphology to the modern human morphology. Large, bony eye sockets, a large nasal opening, and stout facial bones dominate the face of the pongid. The pongid facial pattern is seen in all fossil hominids except anatomically modern man.
![]()
![]()
![]()
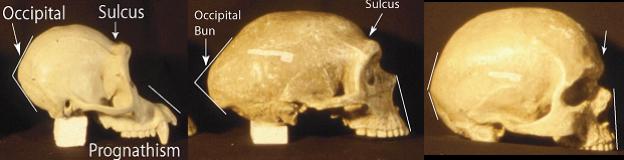
Lateral View. A similar pattern is seen in lateral view. The pongid skull is characterized by a sulcus behind the brow ridges, prognathism and a protruding occipital region. Anatomically modern man lacks prognathism and brow ridges with the accompanying sulcus, and has a much less pronounced occipital bulge (see photo). The fossil hominids, including the Neanderthals, whose endocranial volume averages more than anatomically modern man, show the pongid pattern.

![]() .
.

1. Chimpanzee 2. Australopithecus africanus 3. “Homo”
habilis 4. Kenyanthropus rudolfensis 5-6. Pithecanthropus erectus 7-8. P. soloensis 9. Homo heidelburgensis 10-11. H. neanderthalensis
12. H. sapiens
When viewed from the top, all of the skulls except that of anatomically modern man show a marked constriction just behind the orbital region. This constriction imparts a definite pear-shape to the braincase. Because the skull of anatomically modern man is inflated in the frontal area and has gracile orbits, the overall shape of the skull is more ovoid than pear-shaped.

Another character that is unique to anatomically modern man is the parietal bulge. When viewed from behind, the widest point of the skull of anatomically modern man is high, in the parietal region. The widest point of the pongid skull is close to the base of the skull, near the auditory region. All of the fossil hominids, including Neanderthal, have the widest point toward the base of the skull near the auditory region.
1. Chimpanzee 2. Australopithecus africanus 3. “Homo”
habilis 4. Kenyanthropus rudolfensis 5-6. Pithecanthropus erectus 7-8. P. soloensis 9. Homo heidelburgensis 10-11. H. neanderthalensis
12. H. sapiens

Foramen Magnum. In the pongids, the
foramen magnum is located near the back of the skull so the head can be
inclined and pointed forward during quadrapedal locomotion (knuckle-walking).
In anatomically modern man, the foramen magnum is under the skull since the
skull weight is born by the shoulders while walking upright. The fossil
hominids show intermediate placement of the 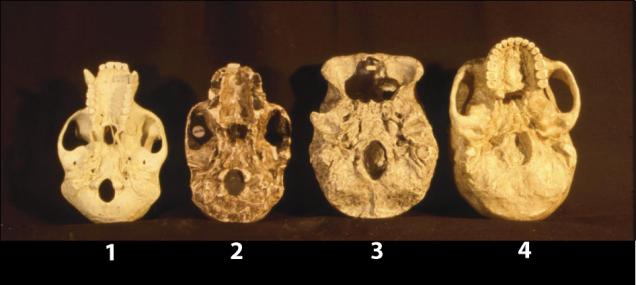 foramen
magnum between that seen in pongids and anatomically modern man.
foramen
magnum between that seen in pongids and anatomically modern man.
![]()

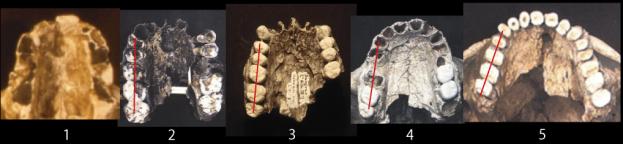
 The
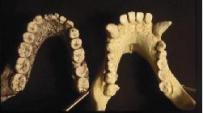
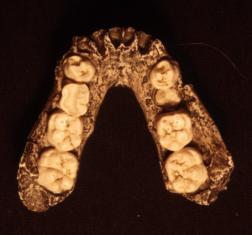
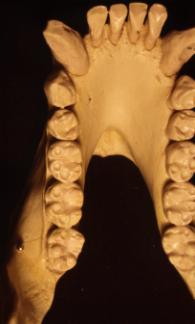
Palate. In the figures above and below, also note the shapes of the palate.
The pongid palate is rectangular with the widest part at the canines then
narrowing backward into the skull. The human palate forms a parabolic curve
with the widest part interior into the skull.
The
Palate. In the figures above and below, also note the shapes of the palate.
The pongid palate is rectangular with the widest part at the canines then
narrowing backward into the skull. The human palate forms a parabolic curve
with the widest part interior into the skull.
The fossil hominid palates are intermediate in shape between the pongid and modern human morphologies. In all of the species except anatomically modern man, the premolars and molars form a relatively straight line. The axis of the line shifts from nearly parallel with the midline of the palate in the australopithecines to broadly divergent in the Neanderthals. Only the Mt. Carmel, Israel, Skul specimens, sometimes placed within the Neanderthals show the parabolic curvature of anatomically modern man.
![]()
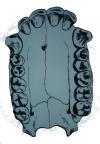
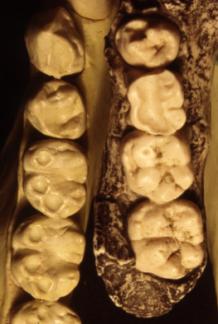
The Mandible. The mandibles of the fossil hominids also share a number of structures with the pongids. The mandible of anatomically modern man has a well-developed protuberance called the mental eminence or “chin”. The pongid mandible lacks the mental eminence and tapers down and back toward the throat. Also seen in the pongid mandible is the “simian shelf” (see figure below), a lingual extension of bone behind the incisors. In the mandible of anatomically modern man, the bone development stops lingually with just enough bone to support the roots of the incisors. All of the fossil hominids including Neanderthal lack the mental eminence and have a simian shelf, although the shelf is relatively short in Neanderthal.
The Teeth. There are also dental differences. In general, pongid teeth are much larger that those of modern man. The canine of the pongids is so large a gap (the diastema) develops between the canine and incisors to make room for the opposing canine so the jaw can close. Modern man has a small canine and lacks the diastema. Except for Ardipithecus and some specimens of Australopithecus afarensis, all of the hominids share the human condition in having a reduced canine and lacking the diastema.
Morphological differences between the teeth also exist. The tooth directly behind the canine in the mandible of anatomically modern man (the p3) is called the “bicuspid”. It has a well-developed metaconid along with the protoconid (see figure below). The pongids have only the protoconid on the p3. The australopithecines have an intermediate morphology with a weakly developed metaconid. If you draw a line through the axis of the p3 and another along the axis of the molar tooth row, the intersection of the two lines is nearly perpendicular in modern man, but forms an oblique angle in the pongids and australopithecines. See the illustration below.
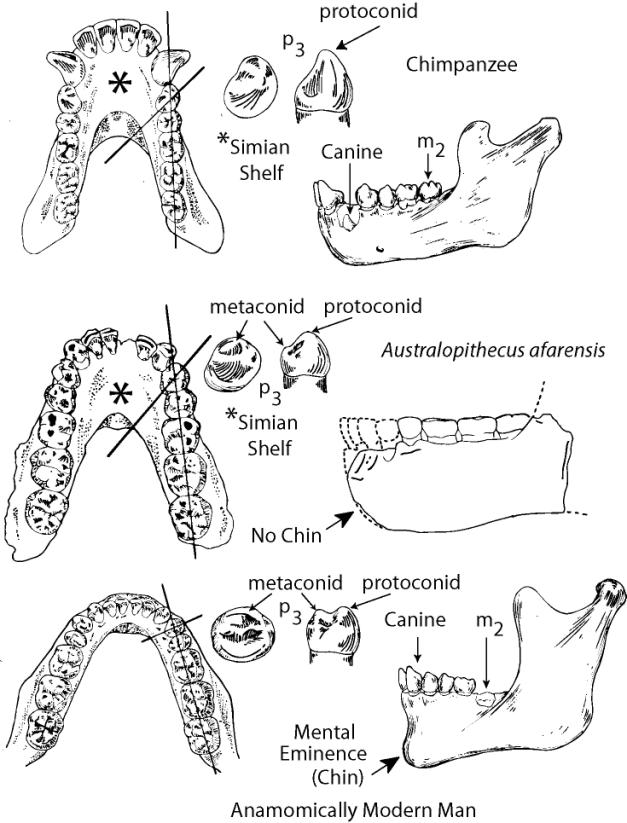
A major physiological difference between humans and apes is the development and emplacement of the teeth. The pongids grow faster and reach maturity earlier than humans. This is reflected in the timing of tooth replacement. Humans have a much longer period of parental care and nursing than the pongids. As a consequence, human infants have a delayed loss of the deciduous premolars (milk teeth). As a result, the adult human canine erupts and is in place before the adult m2. In the chimpanzee, the adult m2 is in place before the adult canine erupts (see figure above). The australopithecines and pithecanthropines show rapid dental growth rates similar to the pongids (Dean et al. 2004).
Data Set II: Hominoid Skeletal Characters
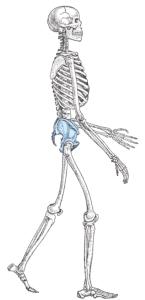
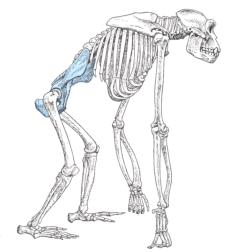
One of the characters used to classify the hominoids in different families is their mode of locomotion. There are distinct skeletal morphologies associated with locomotion. The pongids are tree climbers, but when on the ground, they are quadrapedal knuckle-walkers. The arms are as long or longer than the legs, the scapula has an orientation for supporting the body weight beneath the arms, and the fingers and toes are long and curved for grasping branches. The pelvis is shaped to support the legs and trunk in the bent-over posture necessary for knuckle-walking. Humans are habitual bipeds. The arms are shorter than the legs, the scapula is oriented for holding the arms by the side, and the fingers and toes are shorter and straight. The pelvis is shaped to support the legs and trunk in a vertical position and the knee locks the legs straight to minimize the expenditure of energy when standing. The australopithecines show intermediate character states between pongids and humans, the pithecanthropines intermediate between australopithecines and humans, and the members of the genus Homo are essentially like that of anatomically modern man in skeletal morphology.
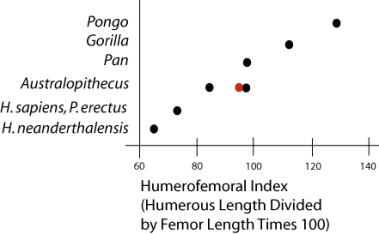
The Limbs. There are a number of indices quantifying the proportional differences in the long bones and trunk of hominoids (Aiello and Dean 1990). As noted above, arboreal apes have longer arms than legs while modern humans have shorter arms than legs. The humerofemoral index quantifies this difference. If the limbs are of equal length, the index will be 100. Values greater than 100 occur when the arms are longer than the legs, values less than 100 occur when the arms are shorter than the legs. The australopithecines plot intermediately between the pongids and humans. The pithecanthropines and Homo species all plot together with the arms shorter than the
legs.
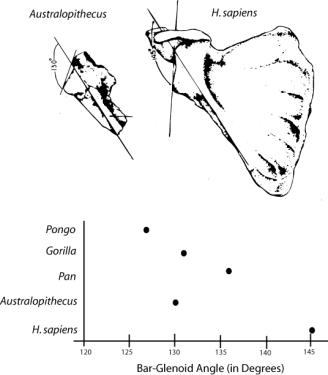
The australopithecines also plot with the apes on the orientation of the scapula, an adaptation for arboriality. If a line is drawn through the axis of the ventral bar on the scapula and another through the glenoid cavity, the intersecting angle can be measured (Stern and Susman 1983). The apes have a bar-glenoid angle less than 140 degrees while humans have an angle of approximately 145 degrees. The australopithecines have an angle about 130 degrees. Like the humerofemoral index, the pithecanthropines and Homo plot together.
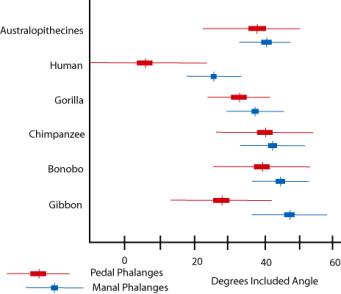
Another index of arboriality is the relative length
and degree of curvature of the phalanges (fingers and toes). The phalanges of
the pongids are relatively long and curved; those of modern man are short and
straight. The australopithecines are short like modern man, but 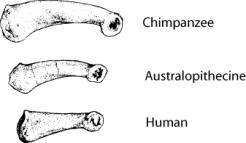 curved
like the pongids. The degree of curvature can actually be calculated and quantitatively
compared among the various hominoids (Susman et al. 1984). The numbers
confirm what was intuitively obvious from the illustration—the
australopithecine phalanges have a pongid curvature. Members of the Homininae
all have relatively straight phalanges.
curved
like the pongids. The degree of curvature can actually be calculated and quantitatively
compared among the various hominoids (Susman et al. 1984). The numbers
confirm what was intuitively obvious from the illustration—the
australopithecine phalanges have a pongid curvature. Members of the Homininae
all have relatively straight phalanges.
 The
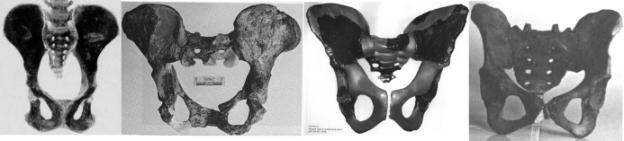
Pelvis. The morphology of the pelvis is a key in determining hominid
locomotor posture. The pongid pelvis is relatively large with an elongate iliac
pillar. The iliac crest is nearly parallel with the back and the angle of
insertion of the femur. The human pelvis is much shorter and bowl-shaped. The
iliac crest is oriented more to the side and forms a much more oblique angle to
the back and insertion angle of the femur. With respect to the angle of the
iliac crest and acetabular fossa, the australopithecine pelvis is intermediate
in shape between the pongids and anatomically modern man. The pithecanthropines
are intermediate between the australopithecines modern man.
The
Pelvis. The morphology of the pelvis is a key in determining hominid
locomotor posture. The pongid pelvis is relatively large with an elongate iliac
pillar. The iliac crest is nearly parallel with the back and the angle of
insertion of the femur. The human pelvis is much shorter and bowl-shaped. The
iliac crest is oriented more to the side and forms a much more oblique angle to
the back and insertion angle of the femur. With respect to the angle of the
iliac crest and acetabular fossa, the australopithecine pelvis is intermediate
in shape between the pongids and anatomically modern man. The pithecanthropines
are intermediate between the australopithecines modern man.
![]()
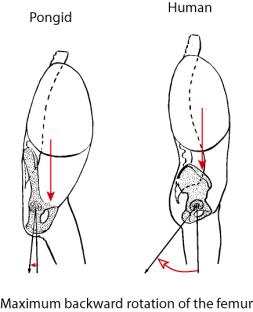
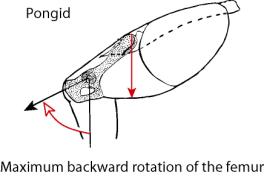 Particularly important for posture are the vectors
for support of the trunk and attachment of the legs. In the bent-over position
of the trunk during quadrapedal locomotion, the elongate iliac pillars of the
pongid pelvis support the back and abdominal tissues (at the expense of torsion
ability). The angle of insertion of the femur allows considerable backward
movement for stride length. When the pongid walks bipedally, the vectors
change. The insertion angle of the femur requires that in order to stand, most
of the backward movement of the leg has already taken place, meaning the that
most of the stride must occur in front of the body axis. The human pelvic
morphology has the vectors optimized for a vertical stance. The pelvis becomes
a bowl to aid in visceral support, the shortened iliac pillar allows for
lateral rotation or torsion of the trunk, and the insertion angle of the femur
allows greater backward movement of the leg, increasing stride and balance when
in the upright posture. The pelvis of all of the hominids has a shortened iliac
pillar more like that of anatomically modern man. The intermediate angles for
femur insertion seen in some hominids would restrict to various degrees the
amount of backward movement of the femur, limiting stride and balance
accordingly.
Particularly important for posture are the vectors
for support of the trunk and attachment of the legs. In the bent-over position
of the trunk during quadrapedal locomotion, the elongate iliac pillars of the
pongid pelvis support the back and abdominal tissues (at the expense of torsion
ability). The angle of insertion of the femur allows considerable backward
movement for stride length. When the pongid walks bipedally, the vectors
change. The insertion angle of the femur requires that in order to stand, most
of the backward movement of the leg has already taken place, meaning the that
most of the stride must occur in front of the body axis. The human pelvic
morphology has the vectors optimized for a vertical stance. The pelvis becomes
a bowl to aid in visceral support, the shortened iliac pillar allows for
lateral rotation or torsion of the trunk, and the insertion angle of the femur
allows greater backward movement of the leg, increasing stride and balance when
in the upright posture. The pelvis of all of the hominids has a shortened iliac
pillar more like that of anatomically modern man. The intermediate angles for
femur insertion seen in some hominids would restrict to various degrees the
amount of backward movement of the femur, limiting stride and balance
accordingly.
Data Summary
There is a gradation in cranial morphology between the pongid skull and anatomically modern man. The australopithecines have a very pongid-like skull; the pithecanthropines are intermediate between the chimpanzee and modern man; the Homo species, while they have a cranial volume as large or larger than modern man, also share many pongid-like characters. Post cranially, the picture is a little different. The australopithecines are intermediate in morphology between anatomically modern man and the pongids, the pithecanthropines are intermediate between the australopithecines and modern man, and there is little difference between of other hominids and modern man.
We will now look at the various hypotheses that have been postulated to explain the data from the three dominant cosmologies: evolution/theistic evolution, progressive creation and young earth creation.
Interpretation I: Evolution/Theistic Evolution
The Model
Through geologic time, mutations and natural selection will produce speciation events. A series of these events will lead from a common ancestor of the pongids and hominids to anatomically modern man.
The Evidence
A number of fossil hominid species have been described. Altogether, these species form a morphological gradient from a chimp-like species to anatomically modern man. The taxa appear in the fossil record in approximately the order expected if they had evolved.
Tests
If evolution occurred, the not only should the species appear in the correct order, but so should all newly evolved characters. Protein and DNA studies show the chimpanzee and man to be more similar to each other than either is to the gorilla, and the African apes and man are more similar to each other than to the Asian orangutan (Jones, Martin and Pilbeam 1992). Therefore, the hominids and the chimpanzee must share a common ancestor to the exclusion of the gorilla. The common ancestor of the chimpanzee and man cannot have any of the character states that evolved later as the two lineages diverged. Any characters shared by fossil hominids and the chimpanzee should have been found in the common ancestor of both. Testable hypotheses can then be built by analyzing the order of appearance of new characters.
The three oldest “hominids” Orrorin tugenensis, Ardipithecus ramidus, and Australopithecus anamensis all lack pelvic girdle and femoral specimens. Whether they were bipedal, and hence truly hominids, remains unknown. They are classified as hominids because the share certain dental characteristics with later species that were demonstrably bipedal. Ardipithecus ramidus is the most chimp-like of the fossil hominids. Orrorin tugenensis is older, but of doubted hominid affinity (Aiello and Collard 2001, Haile-Selassie 2001). All are too incomplete to use for broad comparisons. They share a number of characters with the chimpanzee including cusp morphology of the molars and curvature of the arm and fingers. They also share a number of characters with Australopithecus afarensis (Haile-Sedlassie 2001). Studies comparing them with later hominids have not yet been completed.
Australopithecus afarensis is the earliest hominid well known enough to draw phylogenetic conclusions. Because of this, it is usually placed as the basal taxon from which all other species are derived. The most common evolutionary hypothesis is that A. afarensis gave rise to A. africanus, then “Homo” habilis then to the pithecanthropines (Tattersall 1993). The characters in the first table below tend to support that hypothesis, but the characters in the second table falsify the hypothesis. The actual ancestor to “Homo” habilis would have to be like Australopithecus africanus but have prominent or moderate hollowing above and behind the mental foramen, have the height of the origin of the masseter muscle low, have the height of mandibular ramus relative to breadth low, have the most common site of anterior origin of zygomatic arch nearer the M1, and have rare or absent anterior pillars. There is no known species with all the right character states in the right order (Skelton, McHenry and Drawhorn 1986). When you do this same procedure with all of the other hypotheses of ancestor/descendent relationships, the results are all the same -- there are no unfalsified hypotheses. In practice, the hypothesis that has been falsified the least number of times is accepted as being the closest to what is thought to have actually happened under the principle of parsimony.
|
||||||||||||||||||||||||||||||||||||||||
|
Character States
That Falsify the Hypothesis |
|||
|
Trait |
A. afarensis |
A. africanus |
"H". habilis |
|
Hollowing above and behind mental foramen |
prominent |
reduced |
moderate |
|
Height of masseter origin |
very low |
intermediate |
low |
|
Height of mandibular ramus relative to breadth |
low |
intermediate |
low |
|
Most common site of anterior origin of zygomatic
arch |
M1/P4 |
P4 |
M1 |
|
Anterior pillars |
absent |
present |
rare |
This does not prove that evolution did not occur. You can always invoke the ad hoc postulate that the fossil record is incomplete and if it was complete, the actual ancestors and descendents would be known. Being ad hoc, it is procedurally the same as creationists saying that God did something. Faith is exercised in believing that evolution occurred in spite of there being no unfalsified hypotheses of descent.
The postulate of the incompleteness of the fossil record could be addressed. On the basis of probability, if the fossil record is incomplete by some factor, say 90% for example, and we go to different sedimentary basins, like Laetoli, Hadar, and Chad for example, then, by chance alone, we would have a 90% probability that each species recovered would be previously unknown. In fact, we find most of the species to be the same from these sites even though separated by hundreds of miles. All three basins have yielded specimens of Australopithecus afarensis as well as sharing most of the other mammal species (White, et al., 1984, Brunet, et al., 1995) The percentage of the fauna shared would give a minimum estimate of the completeness of the fossil record. For the sake of argument, say the three sites shared 60% of their fauna. Then the fossil record should not be less than 60% complete. It could actually be more complete if the missing or different taxa were different for some reason other than the completeness of the fossil record, such as biogeography. Whatever the actual calculated completeness turns out to be, that percentage of actual ancestor/descendent relationships should be recovered. The fact that none of the actual ancestors have been identified to date suggests that the ad hoc invocation of the “incompleteness of the fossil record” will continue as will the need to exercise faith that man evolved.
Interpretation II: Young-Earth Creation
The Published Model
Virtually all young-earth creationist models classify the australopithesicenes and “Homo” habilis as extinct apes (Lubenow, 1992, p. 166 and ch. 15) and the other hominid species as degenerate/diseased humans; they postulate that there are no transitions between the apes and modern man (Coffin, 1968; Lubenow, 1992; Marsh, 1958; Maynard-Smith, 1996).
Tests
Are Australopithecines Simply Extinct Apes?
The apes have traditionally been classified by their mode of locomotion. The lesser apes are brachiators, the greater apes are knuckle-walkers and the family of man is bipedal. The australopithecines are also bipedal. To classify the australopithecines with the apes requires redefinition of ape taxonomy. The australopithecines cannot be dismissed by simply changing the definition of what an ape is. They exhibit a combination of human and ape characteristics.

Are There Morphological Transitions Between the Apes and Man?
As noted above, there is a gradation of endocranial volumes from chimpanzee to human.
There is also a gradation in overall morphology. The evidence presented above falsifies the assertion of two distinct (ape and human) morphologies as presented by Lubenow and others.
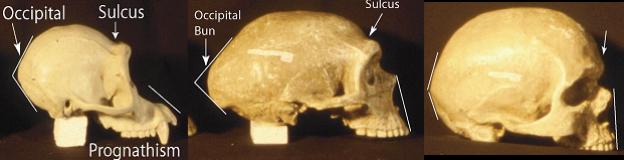
![]()
![]()
![]()
Is the Morphological Gradient the Result of Disease/Degeneration?
Many of the characters found in fossil hominids are also found in pongids, but not in anatomically modern man. No disease stretches the skull and face to produce prognathism and an occipital bun, removes the forehead and creates a sulcus, or creates large eyes with heavy, bony ridges. No disease takes away the chin and adds a simian shelf. These are all characters found in pongids but not in anatomically modern man. In short, there is no “ape character” disease.

![]()
Results of the Test
The hypothesis that there are no morphological transitions between the apes and man has been falsified. The hypothesis that the fossil hominids are diseased humans is also falsified.
Interpretation III: Hybridization
The Model
Another “young-earth” model capable of explaining fossil hominids is that of multiple hybridizations between the pongids, humans and the results of previous crosses. Australopithecus could be the F1 of a human-chimpanzee cross; Paranthropus could be the F1 of a gorilla-human cross. Pithecanthropus could be the F2 from an F1-human cross. F3, F4, etc., backcrosses with humans could produce the other fossil Homo taxa.
The Evidence
Hybridization in an evolutionary model, called reticulate evolution, has been proposed for other, non-primate taxa (Sylvester-Bradley 1979). This is used to explain the blend of characteristics seen in some species. The pattern of blending of character states is called a mosaic pattern, or mosaicism.
The pattern of mosaicism of pongid and anatomically modern human characters seen in the fossil hominids is reminiscent of the pattern seen in other hybrids such as the mule. The mule has some donkey characters such as the external ear shape, some horse characters such as the shape of the skull, and some intermediate characters such as size. The mule is sterile, but other horse crosses such as the zebra/horse cross are not. It also makes a difference which parent is male or female. A male donkey crossed with a female horse produces a mule, which is very different than the offspring of the reverse cross, known as a jenny, which is much smaller and more donkey-like than the mule.

Much of the mosaicism discussed in the earlier sections above has centered on shared chimpanzee-human characters. The genus Paranthropus illustrates shared gorilla-human characters. Paranthropus shares the flat face and sagittal crest with the gorilla, but has a larger endocranial volume and was bipedal (Robinson 1972).
Paranthropus
Paranthropus
The teeth of Paranthropus are very gorilla-like,
while many features of the jaw are intermediate. There is a simian shelf, but
it is much small than the gorilla. The tooth row is linear, but at an oblique
angle to the incisors rather than perpendicular. The incisors form more of a
parabolic arch than those of the gorilla.
In summary, hybridization explains the mosaicism of pongid, human and intermediate character states seen in the fossil hominids.
Tests
It would be technologically feasible perform such a cross. It would also be immoral. At what point in the crosses would “humanness” be maintained? This test should not be done.
While the mosaicism of characters states found within the fossil hominids may be explained by hybridization, hybridization does not easily explain the stratigraphic distribution of the fossil hominids. In general, the australopithecines appear first, followed by the paranthropines and pithecanthropines, then later the species of the genus Homo. There is no evidence of either modern pongids or modern man prior to the appearance of the fossil hominids. (Although there is a fossil orangutan from the Miocene of Pakistan, Sivapithicus.)
The fossil hominids other than Homo appear to have been living and buried as members of a biological community. Their bones are mixed together with other members of the fauna. Their biostratigraphy mirrors the biostratigraphy of their communities.
It is a truism of the fossil record that the higher in the stratigraphic record you go, the more modern-looking the communities become. Young-earth creationists (neodiluvialists) explain this observation through the model of Biome Succession (Ecological Zonation of Clark, 1949). Modern man would have been living at the highest elevations before the flood and would be buried at the highest stratigraphic level. Presumably, this would apply to the fossil record of fossil hominids as well. Biome Succession, however, is just a model, and since there is no direct evidence of modern man before the late Pleistocene, it must be accepted by faith that they could have produced the hybrids postulated to explain the hominid fossil record (or that portion of the fossil record is post-flood and there is some other as yet undescribed process for superpositional community replacement after the flood).
Interpretation IV: Progressive Creation
The Models
There are almost as many variations of “progressive creation” as there are expositors. Most, however, fit into one of four categories: gap theory, day-age, intermittent-day and literary device (Newman 1999). They all accept the secular chronology for the age of the universe and earth and the geologic column as being formed essentially as secular geologists portray it. The gap theory postulates that God originally created the universe including the earth over long ages. All life was later destroyed on earth and the Genesis account is the record of God restoring life on earth about 6000 years ago. The day-age thesis is that each day in Genesis corresponds to a long geologic age of the earth’s history. According to the intermittent-day model, each day of the Genesis creation week is a literal day, but each day is separated by long ages. Finally, there are those who believe that the days of Genesis are a literary device and have no basis in time.
Tests
Conclusions
The exercise of faith is required for any model explaining fossil hominids. For evolution/theistic evolution faith is required in the belief that the fossil record is seriously incomplete and the someday the actual ancestral species will be discovered resulting in an unfalsified hypothesis of descent. In the meantime, faith is exercised to believe the hominids evolved even though no actual ancestor/descendent relationships can be identified. For progressive creation, faith is required to believe that God continually steps in to create new taxa even though the Bible only mentions Creation Week. For young-earth creation, faith is required to believe that man existed throughout the formation of the geologic column even though there is no evidence of anatomically modern man prior to the Pleistocene.
Literature Cited
Aiello, L. and M. Collard. 2001. Our newest oldest ancestor? Nature 410:526-527.
Aiello, L. and C. Dean. 1990. An Introduction to Human Evolutionary Anatomy. Academic Press, New York, NY. pp. 596.
Brunet, M., A. Beauvilain, Y. Coppens, E. Heintz, A. Moutaye, and D. Pilbeam. 1995. The first australopithecine 2,500 kilometres west of the Rift Valley (Chad). Nature 378:273-275.
Clark 1949
Coffin, 1968
Dean, C. M. Leakey, D. Reid, F. Schrenk, G. Schwartz, C. Stringer, and A. Walker. 2001. Growth processes in teeth distinguish modern humans from Homo erectus and earlier hominins. Nature 414:628-631.
Haile-Selassie, Y. 2001. Late Miocene hominids from the Middle Awash, Ethiopia. Nature 412:178-181.
Heinzelin, J. de, J. D. Clark, T. White, W. Hart, P. Renne, G. WorldeGabriel, Y. Beyene, and E. Vrba. 1999. Environment and behavior of 2.5 million-year-old Bouri hominids. Science 284:625-629.
Jones, S., R. Martin and D. Pilbeam. 1992. The Cambridge Encyclopedia of Human Evolution. Cambridge University Press, Cambridge, UK. pp. 506.
Leakey, M. et al. 1996. A record of faunal change in the late Miocene of East Africa. Journal of Vertebrate Paleontology 16:556-570.
Lubenow, M. 1992. Bones of Contention: A Creationist Assessment of Human Fossils. Baker Books, Grand Rapids, MI. pp. 295
Maglio and Cooke 1978
Marsh, 1958
Maynard-Smith, 1996
McDougall, I. and C. Feibel. 1999. Numerical age control for the Miocene-Pliocene succession at Lothagam, a hominid-bearing sequence in the northern Kenya Rift. Journal of the Geological Society of London 731-745.
Pickford, M. and B. Senut. 2001The geological and fauna context of Late Miocene hominid remains from Lukeino, Kenya. C. R. Acad. Sci. Ser. Iia 332:145-152.
Robinson, J. 1972. Early Homind Posture and Locomotion. Univ ersity of Chicago Press, Chicago, IL. pp. 361
Skelton, R., H. McHenry and G. Drawhorn. 1986. Phylogenetic analysis of early hominids, Current Anthropology 27:21-35.
Stern, J. and R. Susman. 1983. The locomotor anatomy of Australopithecus afarensis, American Journal of Physical Anthropology 60:279-317.
Sylvester-Bradley 1979.
Susman, R., J. Stern, Jr., and W. Jungers. 1984. Arboreality and bipediality in the Hadar hominids. Folia Primatologica 43:113-156.
Tattersall, I. 1993. The Human Odyssey: Four Million Years of Human Evolution. Prentice Hall, New York, NY. pp. 191
White, T., R. Moore, and G. Suwa. 1984. Hadar biostgratigraphy and hominid evolution. Journal of Vertebrate Paleontology 4:575-583.
WoldeGabriel, G., et al. 2001. Geology and paleontology of the Late Miocene Middle Awash valley, Afar rift, Ethiopia. Nature 412:175-178.
Wood, B. and M. Collard. 1999. The human genus. Science 284:65-71.