The Origin of Animal Body Plans. Wallace Arthur, Cambridge 1997. Notes:
Page
x "It probes the question of how the morphological ‘designs’ of the thirty-five of so animal phyla arose in a burst of creative evolutionary activity in the distant geological past."
xi "New syntheses between established disciplines are catalyzed by individuals adopting uncomfortable positions at interfaces."
7. "Regardless of the timing of early divergences, it appears that no phylum-level body plans have arisen in the animal kingdom in the last 500 my."
8. "… the main message to emerge is twofold: (a) animal evolution was more ‘creative’ before 500 my ago than since then, when it has been rather conservative, so there is a bread parallel with development, as suggested at the outset; but (b) there is considerable uncertainty over the number, nature and timing of early creative ‘explosions’."
- "In fact, Neodarwinian theory is incomplete even when assessed against its own internal criteria. The main problem with neodarwinism in its current form is that theoretical structure is extremely lopsided. There is nothing wrong with elaborate models of selection, but a detailed quantitative statement of how existing types are sorted and selectively eliminated (or held in a state of stable equilibrium) cannot pretend to be a complete theory of evolution.
11 The only approach to evolution that has attempted to focus on creative forces has been that of Evolutionary Developmental Biology.
12 "Systematists focus their interest on elucidating, and interpreting what can be variously called Nature’s hierarchy (Rosen 1984), the natural hierarchy, or the pattern of natural classification (Panchen 1992). This is the pattern of nested homologies that we see in the animal kingdom (and also in plants, where it is partially observed by frequent occurrences of reticulate evolution (Grant 1981), and which Scotland (1992) includes among the axioms of cladistics….characters involved in the "outer" homology of greater taxonomic breadth, also appear earlier in development (ref)…..there is no reason why such a pattern should have been predicted and indeed even in retrospect there is no way that it can be explained without bringing in some additional information---" He then proposes that he can bridge this unhappy situation by making the early genes more conserved (since they are more deleterious to the organism).
- "There is, however, a problem. Those genes that control key early developmental processes are involved in the establishment of the basic body plan. Mutations in these genes will usually be extremely disadvantaged, and it is conceivable that they are always so […] But it is clear, for example for cross-taxon comparisons of a particular group of such genes—the homeobox—containing Hox genes (ref) (p.15) that the homologous developmental genes in different phyla are different. Even where there is an unexpectedly high degree of similarity (e.g. 98 percent between some human and Drosophila homeoboxes—Boncinelli et al. 1985), there is still a difference. The genes concerned have evolved. And…early developmental pathways have changed in consequence. Does this mean that mutations of these genes are sometimes advantageous? If so are these in some sense successful macromutations? There is a can of worms here which has been opened with generally negative consequences on several occasions, most notably by Goldschmidt (1940). (p.16) But we cannot bypass this issue, however unpopular."
He approaches the question of defining the term "macromutation", a term over which there appears to be little consensus, with appropriate caution, suggesting the concept is flawed since all mutations can be arranged on a continuum based upon the magnitude of their effect. He then demonstrates by analogy how early-developmental mutations can produce quantitatively larger effects than later ones. He then presents one of the major theses of the book, a concept intuitively obvious but not previously stated:
22 "While in a non-developmental explicit approach it seems plausible that many small changes can indeed accumulate to give a larger one, in a developmentally explicit approach it is clear that many late changes cannot accumulate to give an early one." Thus if taxonomically distinct [human and Drosophila, for example] organisms differ right back to their early embryogenesis, as is often the case, the mutations involved in their evolutionary divergence did not involve the same genes as those involved in the typical speciation event, where usually the early embryogenesis of the daughter species concerned are virtually identical."
"What is a body plan? It is easier to give examples than to define, but clearly includes a group of morphological characteristics that define a class of organisms. We speak of the gastropod body plan. After studying a single example, one can recognize another organism with the same body plan." Having established the concept, Arthur considers how it might be used in evolutionary analysis. He considers four methods that have been used to analyze morphological features as primitive or derived: the commonality principle, stratigraphy, the outgroup method and the ontogenetic method. The first states that features shared by more organisms will generally be more primitive (earlier evolutionary origin) than those that are less common. An example will suffice to illustrate the difficulties with this approach. In insects wingless condition is considered primitive. Yet winglessness is present in only a few orders, and an analysis based upon the commonality of wings would result in very strange evolutionary conclusions.
- Macro vs. micro – Developmentally early mutations have large consequences; developmentally late mutations have smaller consequences. We cannot, by developing lots of late mutations, effect a change in a developmentally early gene. Therefore, meaningful mutations in terms of phyletic events will always be macromutations.
- "First, the body plan that comes to mind for a particular species is invariably one which characterizes a higher taxon, but the exact level varies."
- "However, the central point remains the same: characters used in the delineation of body plans are major features of the overall layout of the adult body of the developmental trajectory giving rise to that adult body."
- "Ironically, the concept of a body plan also becomes less useful above the level of the phylum."
"… interrelationships between phyla are usually obscure (ref)…"
- "Estabrook make the point as follows: ‘the relatively more primitive states are likely to be distributed more generally throughout the group under study’."
"Although there are some cases in which this approach works, it has been heavily criticized…"
AT WHAT LEVEL DO INSECTS FIRST APPEAR IN RECORD?
- "While the use of ‘commonality’ is flawed in principle, the use of stratigraphic information can be troublesome for practical reasons – stemming from the incompleteness of the fossil record.
"Although there are some cases in which reality comes tolerably close to this ideal situation [complete record = perfect phylogeny], they do not include body-plan origins, where stratigraphic information is usually very limited (ref)."
"… on its own it cannot be expected to enable us to make sound decisions on which character states are primitive and which derived."
- "Unfortunately, different characters often suggest different systematic patterns as a result of convergent evolution, and the initial naïve expectation that molecular data would be sufficiently free from this problem to render phylogenies ‘obvious’ has been shown to be unwarranted ." (QRB 64: 291-318)
"Here, we provisionally accept whichever cladogram minimizes the amount of convergence that we are forced to postulate. This can be a highly complex matter when many characters are involved – and indeed, there are different kinds of parsimony…"
- "Outgrouping assumes relationships are known then infers characteristics of the outgroup has in common with the in-group are primitive. Unfortunately different characters often suggest different systematic patterns as a result of convergent evolution, and the initial naïve expectation that molecular data would be sufficiently free from this problem to render phylogenies ‘obvious’ has been shown to be unwarranted (ref)."
- Principle of parsimony is then invoked: we provisionally accept whichever cladogram minimizes the amount of convergence that we are forced to postulate.
Cladistics is the death knell of evolution—one convergence postulated may be sufficiently unlikely to falsify the theory.
- "The cases in which we have a clear and agreed picture of the nesting of Unterbaupläne within phylum–level body plans, and of the relationship of those major body plans to each other, still represent a small fraction of the overall pattern, …"
41. Finally, ontogenetic analysis can only be expected to be useful if the characteristics being studied are those "late developmentally…" "Current neo-Darwinism permits virtually any pattern, which is one of the criticisms that some cladists (such as Rosen 1984) level against it." Genes that regulate development and shape etc. Who even says such genes could even exist, yet they were present before the first organism preserved as a fossil.
43 First, what overall form does the pattern of interaction between developmental genes take?…feedback loops…crosslinking…redundancy…All of this argues for the existence of a highly complex network. However, which this may seem like a recipe for despair….all of this of course has major evolutionary implications.
- "The high degree of repeatability of ontogeny implies some sort of redundancy… All of this argues for the existence of a highly complex network. However, while this may seem a recipe for despair, the network is likely not to be completely irregular; rather it will be patterned in various ways which we must seek to describe."
"I must admit that I was initially of the view that the ‘chromosomal layout’ aspect of genetic architecture was generally unimportant, but the discovery of ‘colinearity’ has made that view untenable (ref)… These observations are clearly telling us something, even if it is not yet clear what."
- "With regard to the origin of the major (phylum-level) body plans, one thing is clear: they do not result from invasion of the different major classes of environment: land, sea, freshwater and air. All of the major animal body plans are marine in origin."
- Let us now turn to maintenance of body plans…All characters originate, but most also disappear again or are fundamentally transformed; that is, they are evolutionarily transient. One of the main features of body plans, in contrast, is their high degree of conservation, extending over hundreds of millions of years and – in the main phyla – across thousands of species."
What causes this extraordinary degree of conservation??? "…developmental constraint…"
It seems in reading through this section that he has considered every possibility but the correct one.
(p.53) Molecules, like morphologies vary in their evolutionary rates and are subject to parallel and convergent evolution; and in consequence different molecules often suggest different phylogenies, just as do different morphological characters.
(p.59) The reality of a deuterostome grouping of chordates, chordates and echinoderms is naturally unchallenged. […] The evidence for a monophlyetic protostome clade is less convincing…
(p.63) In the view of some ‘transformed’ or ‘pattern’ cladists, even the connection with phylogeny [in cladograms] is unnecessary (Plantics, 1979), evolutionary lineage divergence being regarded only as a possible explanation of the pattern observed in the cladogram, and one that should not be assumed at the outset: despite the difficulty of imagining an alternative explanation.
(p.64) From a body—plan perspective, the most pressing task is to attempt to date the origins of the various annual phyla…(p.65) the following groups were present in the early Cambrian: Porifera, Cnidaria, mollusca, Annelida, Crustacea, Chelicerata, Brachiopoda, Echinodermata…Chordata…Hemichordata…all but one of the major radiations of phyla…had already occurred by this time. The one possible exception is the Aschelmenthes, but these have a very poor fossil record…and…may well have been present….
(p.93) Gould’s Ontogeny and Phylogeny (1977a) is partly and exposition of the importance of heterochrony in evolution. While I agree that it is indeed important, I doubt if heterochrony is the key to understanding the origin of body plans—rather it relates to the proliferation of variants of these plans. New structures and organs have to be produced before they can be accelerated or retarded developmentally in relation to each other. (p.94) …"No general theory of development has emerged in spite of the mounting mass of observational and experimental information"…as true now as it was [in 1961]. Because we have no theory, we cannot be certain that one is possible (or that it is not).
In contrast, Needham (1934, p.221) is of the view that "Embryology in particular has been a theoretically threadbare, since the decay of the evolution theory as a mode of explanation…
(p.107) make sure Cadhedrin is covered on conserved slide
(p.110) Talks about coevolution in receptor ligand systems ---hilarious—what is the mechanism since any change in one automatically makes the organism less fit and it would be eliminated—these would be macromutations—makes no sense, but is necessary to postulate—of course this begs the question of why they are so similar if they do evolve this way…
Sonic hedgehog genes are responsible for many developmental failures welding innervation and limb bud differentiation in insects and man—brain development, feathers in birds and insect eyes.
(p.118) "As might be expected, many aspects of the ecdysone gene switching system…are evolutionarily conserved [over 100 genes] between species…families…and possibly also between orders and classes."
(p.120 para5.6)A key issue is raised: focus on one event or production of one hormone w/o following precedent cascade of events leading up to it—infinite regression—
(p.123 para2) if a name can be given to something too complex to comprehend it somehow gives us the feeling that we understand it—
(p.126) "…the number of genes involved in development has turned out o be larger than was once anticipated."
(p.137) "One thing that is apparent, however, is that colinearity is not a chance phenomenon—the odds against that are too great."
(p.141) "There is not one but at least six Ubx products—isoforms of the protein produced by different RNA splicing patterns…"
(p.143) "The reasons for this colinearity are not yet apparent, but the probability of it being due to chance is sufficiently low that it can be regarded as negligible. Given the antiquity of the homeotic gene complex(es), and the consequent opportunity for ‘reshuffling’, there is clearly a selective reason for the maintenance of colinearity….(Despite this, it has been shown that at least some aspects of homeotic gene function are insensitive to chromosomal rearrangements:[ref]."
(p.146,7) Development works like a computer with at least three instructions: Do-stop-and wait. And all of the programs "…are highly complex, involving an interplay between multiple types of signal in a network that includes adversity of feedback loops."
"Turning now to evolutionary theory, it is ironic that the sheer complexity of interaction between genes that underlies development carries a very simple and clear evolutionary message […]’…interval selection…"[that is the genes must be responding not to external pressures of temperature etc, but to changes in other genes with which they are associating].’
150. Results of work:
- confirmation of animal monophyly and the strong conservation of developmental genes
- …
- …
(p.150) "…One of the main aims of evolutionary developmental biology…is to map developmental genetic control systems onto the phylogeny of descriptive embryologies that we already have for the animal kingdom.[…] three robust messages deriving from this accumulating body of work are: conformation of annual monophyly and the strongly conserved nature of developmental genes…; realization of just how important gene duplication events were…; and appreciation of the ubiquitous evolutionary importance of changes in interactional architecture and spatiotemporal expression patterns.
(p.152) "(Studies on Ubx in Drosophila suggest that the number of downstream targets for that single Hox gene may be as great as 170….)"
"The brood category of homeobox-containing genes extends beyond the annual kingdom into the other main eukaryotic grouping (plants and fungi…). However Hox genes, as a subset within this broader category, appear to be restricted to, and distributed throughout, the animalia.
-"no Hox genes in protozoans… the most ‘primitive’ and phyla such as Porifera and Cnidara do contain Hox genes…
(p.155)-"It appears that an original cluster of thirteen genes replicated to produce four such clusters, and that different secondary losses in different clusters gave rise to the thirty-nine genes found in today’s higher vertebrates.
-orthology = cross-species homology
-parology = cross-locus homology w/u species
(p.156) "Maintenance of Hox genes in clusters, given the time available in which could have disintegrated and the genes in them dispersed, must be governed by selection….However this selection appeared to be relatively weak in drosophilids…"
(p.165) "The ad hoe nature of evolution must always be borne in mind."
(p.166) "Yet while this type of transition [protochordate to tetrapod] was associated with an elaboration of Hox genes in chordates, it is not associated with a comparable elaboration in the annelid/arthropod clade." […] Perhaps the move from a simpler, non-segmented protostome body plan to proto-annelid was accompanied by Hox gene proliferation. […] However […] the apparent existence of up to ten Hox genes in flatworms is hard to explain under this view.
(p.168) "Colinearity [spatial as well as temporal order of activation] is maintained in transgenic mouse embryos containing regulatory elements from Drosophila"
- "Colinearity still has much to teach us, both about development itself and about its evolution."
(p.169,70)Allows that natural pattern "may be as valued a way of viewing these things as "evolutionary." (p.172) Also, homology has a non-evolutionary origin. "Despite the widespread acceptance and usage of the concept of homology, its familiarity to evolutionary biologists tends to mask considerable problems associated with it"
(p.172) "Some cladists wish to exclude evolution from the definition of homology, so that an explanation of it in evolutionary terms is not ‘circular reasoning’(Brady 1985)." In that case homology is hard to define. "[…]" My own view is that it is perfectly acceptable to define homology in evolutionary terms[…] What is difficult[…] is being certain that a particular pair or group of characters from different taxa, are not homologous.
homoplasty- convergent
- "Given this proposed homoplastic relationship between insect and vertebrate limbs, we would not expect their ‘detailed complexities’ to be similar…"
(p.174) "…When the protostome and deuterostome branches of the bilatera radiation split more than 500 my ago, their last common ancestor was boneless." Limbs were invented independently by arthropods and chordates. The tetrapod leg originated from a fin (…), and fins in turn originated from relatively insignificant out-growths from the trunks of amphibious like protochordates. The arthropod limb probably arose during the early history of the arthropod clade, so that (for example) insect and crustacean limbs may be homologous (….), just as are mammal and reptile limbs; but the two groups are not homologous to each other. […] Given this situation, it is perhaps odd that some important aspects of the developmental-genetic control of limb development are rather similar between insects and vertebrates. In particular, the hedgehog gene … is expressed in the posterior compartments of wing and leg imaginal discs in Drosophila (ref). Its close homologue, Sonic hedgehog … has a similar pattern of expression in [humans,] the limb buds of mice and chicks (ref) and in the fin buds of zebrafish..."
"If the genetic control of limb formation was entirely distinct from the control of development in the trunk, then these expression patterns of hedgehog family genes would present us with strong evidence that the prevalent view of metazoan phylogeny was wrong, and that arthropod and chordate limbs were homologous. However […] hedgehog family genes are also involved in developmental processes in the trunk. This provides a possible [emphasis added] explanation for developmental-genetic similarity despite homoplasty. […] In reality, the pattern of cellular movements and differentiation involved in the formation of a limb primordium is very complex:[…].
"Clearly this sort of thing could happen twice or many times in phylogenetic history, with the result that homoplasty limbs have similar expression patterns of developmental genes because of their origins from homologous trunks [sic]. Perhaps, then, there is no need to revise our picture of metazoan limb phylogeny after all. (Whether a broadly similar form of argument can be used to defend the previously agreed homoplasty of arthropod and chordate segmentation against recent suggestion of homology based on developmental-genetic similarity (…) remains to be seen."
(P.178) "Very high amino acid sequence similarities are observed among ‘core’ members of the cadherin family. For example, K-cadherin from rats has around 97 percent sequence similarity with human cadherin-6 (…)."
-Cadherens tend to cluster with respect to type then with respect to species, ie:
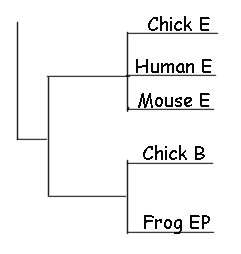
(p.180) "Let us return to the matrix picture…whose columns are either developmental mechanisms or developmental genes, and whose rows are species. If it is indeed true that there is typically less variation going up and down a column than there is in traversing a row, then this argues for the establishment of different types of developmental genes (and mechanism) at a very early stage in metazoan evolution.
(p.182) Gene duplication and mutation-speculation on speculation on speculation
(p.184) "The evidence that many-perhaps even most-genes arose through duplication events is now overwhelming (…)."
(p.191) "The main problem besetting Goldschmidt’s (…) macromutational theory of the evolution of development was the severely detrimental effect of macromutational changes in key developmental genes. However, most known smaller-effect mutations of these genes that have been well studied are also detrimental.
(p.203) "… in general, early acting developmental genes will have more major effects than their later counterparts. This probabilistic trend cannot fail to have evolutionary consequences."
Chap. 9 The spread of variant ontogenies in populations
(p.210) "Ability to spread through populations, on the other hand, most certainly is in doubt for many mutations of developmental genes, … no theory of evolutionary mechanism which fails to address this issue is ever likely to gain widespread acceptance." […] It seems likely that the probability of population-level spread to fixation is zero for many developmental mutations…because of their detrimental effect on internal coordination.
(p.216) He does Hardy Weinberg, but for a dominant mutation! Slipped that one in--"If the new, advantages mutant allele is recessive (which is more commonly the case), then it may be dependent on drift to raise its initial frequency to a level at which homozygotes …begin to appear, because prior to such appearance, selection is unable to act" he concedes-later "In other cases [than dominant], the trajectories are quantitatively altered but qualitatively the same in that fixation still occurs." Give me a break!
Talks about internal vs external selection-everyone has ignored importance of internal selection, which has been raised as an issue for 30 years.
(p.226) "These early animals…consisted of relatively few cells compared with most present-day animals. Their ontogenetic trajectories were therefore much simpler than those to which most of our detailed embryological information relates [wishful thinking]." Also"…their ontogenies were almost certainly more evolutionarily flexible than those of their later counterparts.[…] Mutation of genes controlling early development thus occurred at the beginning of animal evolution [!], in a very different ontogenetic context to those occurring in Caenorhabditis, Drosophila, Mus or any other present-day animal."
(p.229) "It may be, however, that a rather similar dynamics operating on radical developmental variants in primitive small-cell-number [uh-huh] annuals occupying the relatively ‘un-cluttered’. [understatement of the year] niche space of the Vendian (or earlier) could produce much more dramatic results;…."
(p.229 ff) Consider a mutation in a gene in early development in Drosophila-It will scramble normal development-lower coadaptation so organisms either die before adulthood or are unfit to compete. (p.232) "Because of these characteristics, it is unlikely that such mutations make a significant contribution to present-day evolution….And indeed, it seems equally unlikely that they have had a significant role over the majority of animal evolutionary history. However, in the very special conditions…prevailing in early animal evolution, the fate…may have been very different.
Talks about Vendian empty niche space-but there couldn’t have been any with all of those Precambrian species around!.
(p.233) "the scenario pictured is not a highly probable one, but that is not the point."
"The sudden appearance of a morphological novelty [by the highly impossible scenario above] in the manner described above does not, in itself, constitute the origin of any body plan. Body plans incorporate many deep rooted and interlocked features [emphasis added], and the origin of any body plan in inevitably a multistage process."
(p.236) "Once a new body plan has become established…it then becomes stabilized[…] further mutations will tend to damage this new found developmental integration. In some respects [guess which ones], this maintenance of the body plan is of more interest than its origin:[…] all phenotypic characters have an evolutionary origin, but only a very few of them---body-plan characters---are maintained for hundreds of millions of years"
Places his views "somewhere between" non uniformitarian Lamarckism and panselection and conventional uniformitarian views.
Chapter 10 Creation vs destruction.
(p.242) Quotes (negatively from Doner "Mutations with larger phenotypic effects (macromutations) cannot enjoy the services of [positive] natural selection.)
(p.243) With respect to special creation and Lamarkism "Evolutionary theory should be now mature enough to disregard these old adversaries.
[…] the time is ripe for a major drive to increase our understanding of the creative side of evolution[!]…."
Develops concept of morphospace-a hypothetical construct that avoids consideration of mechanism.
(p.254) "The creative side of selection is harder to see than the destructive side of mutation and requires a larger line scale. In the short term, the only possible results of selection…are a) changes in the relative frequencies of preexisting variants and b) the local extinction of one of them."
Chap 11 Ontogeny and Phylogeny Revisited-
Attempts to establish ontogenetic hierarchy parallel to phylogenetic hierarchy.
(p.259) "Lineages that separated earlier in phylogeny might be expected, typically, to be represented today by species whose morphological disparity is greater than that between species whose lineages separated more recently. But as Hennig pointed out this is not necessarily the case; and there are many examples of a lack of morphological (genealogical) correspondence…[origin of birds cited as example]
(p.263) Referring to his model requiring a hierarchy of ontogeny he states "This is simply an act of faith: I cannot believe that such a directional process as ontogeny is underlain by a genetic architecture that has no hierarchical component."
(p.270) "If Body Plans arose through experimental probing of morphospace by ‘radical’ mutations, and were enhanced and stabilized by internal selection (…), then some very early speciation events may have been fundamentally different from any that we can observe today."
"Phylum-level body plans probably arose through such events [speciation], perhaps tightly packed together in relatively short periods of geologic time…."
(p.276) Points out that the Van Baerian patterns of vertebrates are related to the protected developmental environment and organisms developing without this do not follow Van Baer’s "law" ie most organisms.
Chap 12: Prospect: Expanding the Synthesis
Check out Rosen 1984 Hierarchies & History. In Evolutionary theory paths into the Future ed. Pollard, J. W. Wiley Chichester UK
(p.285) "General theories are often built up painstakingly from many different components. They are underpinned by various ‘foundations’ Different parts of a theoretical edifice are often interdependent-if one part turns out to be wrong, the whole structure may eventually collapse."
Points out mild critics of evolution are reacted to with boredom. Radical critics with "heresy". Places himself in between.
Draws a pizza diagram of evolutionary science and contends that about 1/3 is the whole of origins.
(p.289) "Let us now briefly examine some of the evolutionary circle’s deficiencies as a conceptual model.
Check out Gilbert, S. F. Opety. J. M. & Ruff, R. A. Resynthesizing Evolution and Developmental biology. Dev Biol 173, 357-372.
Finishes book with several paragraphs on things that should be done. Find some Precambrian fossils! is one-