
Traduzido por Márcia Oliveira de Paula
Introdução
O mundo atual está na crista da onda do aumento de informações na área de Biologia Molecular. As informações estão sendo geradas tão rapidamente que é impossível que os pesquisadores da área se mantenham atualizados com o fluxo de dados; assim, a interpretação fica atrasada em relação ao surgimento dos dados. Nossa compreensão das complexidades de estrutura e função dentro da célula é revolucionada mais e mais quando descobrimos novos detalhes dos processos celulares. Neste contexto, é uma tragédia particular que nós continuamos, cegamente, dogmatizando uma teoria fundamental da biologia que tem mais de 150 anos: a teoria da evolução.
É compreensível que Darwin e outros dos primeiros protagonistas da teoria da evolução não levassem em conta as dificuldades envolvidas na evolução de formas de vida complexas, quando quase nada se conhecia sobre elas. Este era o caso quando Darwin começou a formalizar sua versão de uma teoria para a evolução espontânea e não dirigida de formas de vida, na primeira metade do século 19. O mesmo aconteceu no século subsequente. Mas, durante os últimos 30 anos, o quadro tem sido mudado pelas ferramentas da sistemática molecular moderna, juntamente com os avanços em nossa compreensão dos processos celulares e moleculares em uma ampla variedade de organismos. Agora é impossível fazer comparações detalhadas dos aspectos moleculares de uma grande variedade de organismos e construir ligações filogenéticas entre estes organismos, baseando-se nestas comparações. Com estas poderosas ferramentas à disposição, não é mais necessário supor que tipos de processos operavam nos organismos extintos, que não estão mais à disposição para serem estudados. Muito da arquitetura molecular de tais formas pode ser reconstruída a partir de dados facilmente disponíveis para nós atualmente. As conclusões de tal trabalho são muito surpreendentes e compreendem o tópico desta apresentação.

Figura 1. O trilobita Devoniano Phacops moroccensis das montanhas do Atlas em Marrocos. Esta amostra espetacular tem características complexas tais como olhos compostos (neste case, esquizocroal), nadadeiras, membros compostos articulados, antenas, forma de corpo cefalizada, partes bucais compostas e outras coisas complexas que são comuns tanto em insetos modernos como nos primeiros trilobitas encontrados no Cambriano inferior (camada geológica mais antiga que tenha fósseis) ao redor do mundo! O que podemos aprender sobre biologia molecular nesta amostra de animal extinto?
Os trilobitas são membros extintos do filo Arthropoda, ao qual pertencem os insetos modernos. Estas criaturas deixaram um registro fóssil longo e detalhado nas rochas, começando no início do Cambriano e terminando no Permiano. Os trilobitas eram formas requintadas, que possuíram corpos segmentados elaborados, com sistema nervoso cefalizado, apêndices torácicos e abdominais articulados, antenas e olhos compostos. Devido ao fato de os trilobitas serem formas extintas, sabemos muito pouco sobre seus hábitos de vida, exceto pelo que podemos deduzir pela sua associação com outras formas que têm representantes vivos, e a partir também de reconstruções cuidadosas dos ambientes deposicionais nos quais eles são encontrados. Entretanto, a teoria da evolução fornece-nos um mecanismo para reconstruir, em detalhes inimagináveis, a fisiologia e a biologia molecular destes primeiros tipos de metazoários, amplamente distribuídos.
Esta reconstrução tem um grande significado em nos fornecer um quadro da riqueza e complexidade das primeiras criaturas metazoárias. Esta reconstrução também contribuirá substancialmente para nossa compreensão dos processos que teriam que ter precedido o aparecimento destas surpreendentes criaturas que, em quase todos os lugares, marcam os limites entre rochas essencialmente desprovidas de vida metazoária e aquelas rochas que contém evidência abundante de vida metazoária. Antes de começarmos a explorar a natureza dos trilobitas, deixe-nos lançar algumas bases fundamentais para as premissas que exploraremos em nossa reconstrução. Eu irei:
- demonstrar, usando a suposição fundamental da teoria da evolução, que podemos conhecer, em detalhes requintados, a biologia molecular do trilobita.
- mostrar que os trilobitas são tão complexos, a nível molecular, como qualquer forma moderna;
e, na ausência de qualquer evidência física da evolução de sistemas complexos, ou do aumento no conteúdo informacional dos sistemas complexos existentes,
- mostrar que a crença na teoria da evolução é uma questão de pura fé, já que não há evidência física para a melhoria no conteúdo informacional de qualquer sistema complexo. Outra teoria que explica as origens – a Criação Especial – tem precedentes científicos, porque ela realmente oferece uma explicação para as origens que se harmoniza com os dados disponíveis.
Origem das Células
Todos os seres vivos, incluindo os trilobitas, são compostos por células. A teoria da evolução propõe que estas células surgiram num passado distante, a partir de um ou mais sistemas viventes complexos, derivados por processos naturais de materiais presentes na terra pré-biótica. Estas proto-células primitivas se estabeleceram e, durante vastos períodos de tempo, desenvolveram sistemas complexos capazes de duplicação eficiente dos componentes necessários para a vida.
Durante este período, os detalhes do código genético foram trabalhados, os sistemas de enzimas necessários para a duplicação do DNA foram aperfeiçoados, as enzimas necessárias para produzir RNA–mensageiros funcionais desenvolvidas, e a maquinaria para produzir proteínas a partir da informação contida no RNA–mensageiro foi estabelecida. Uma área de especulação atual é se este sistema moderno foi o primeiro a se desenvolver ou se um sistema mais simples, envolvendo apenas moléculas de RNA capazes de auto-duplicação e atividade catalítica, o precedeu.
Esta última sugestão apareceu, em primeiro lugar, para oferecer uma solução para o dilema criado pelo requerimento do aparecimento simultâneo de proteínas e DNA para codificar para aquelas mesmas proteínas. Mas atualmente existe pouca evidência de que estas moléculas de RNA catalíticas ou "ribozimas" tenham um papel altamente significativo nas células modernas e permanece enigmático o problema da mudança de um sistema de ribozimas para um sistema de proteínas governadas por DNA. Apesar de a origem da vida não ser o assunto deste trabalho, vale a pena observar que este cenário, ou qualquer outro que explique a origem espontânea de uma célula viva, pertence ao campo da ficção científica. De qualquer forma, é muito evidente que, antes do início do Cambriano, os detalhes das células eucarióticas modernas, das quais os trilobitas eram compostos, estavam totalmente concluídos, como veremos.
Revelando o Passado
O que podemos saber sobre a biologia molecular, celular e fisiologia do trilobita? A premissa fundamental desta apresentação é que podemos determinar, em detalhes preciosos e requintados, os mecanismos que operavam nas células e tecidos dos trilobitas. Esta premissa está baseada em uma síntese fundamental da teoria da evolução: a de que características compartilhadas em comum por organismos distintos, a nível molecular ou celular, são devidos a ambos terem uma ancestralidade evolutiva comum.
Esta suposição é amplamente aceita e envolve toda a teoria da evolução, sendo a base de toda a taxonomia evolutiva moderna. Apesar de algumas similaridades anatômicas serem consideradas exemplos de evolução convergente (derivadas independentemente e não relacionadas geneticamente), tais como as asas de insetos, répteis, aves e mamíferos, tais casos são facilmente identificáveis, e as similaridades que existem a nível celular e molecular são geralmente consideradas como indicadores de ancestralidade comum.
Assim, as características moleculares compartilhadas pelas ervilhas e pelo homem (Fig. 2) requerem que tenha havido, em um passado longínquo, um ancestral comum a ambos, que possuísse estas características comuns. (árvore filogenética, segundo Wray et. al). Qualquer outra conclusão requereria que eventos altamente improváveis tivessem acontecido repetidamente, com exata precisão, e isto falsificaria a suposição fundamental da sistemática molecular e tributaria a credulidade além dos limites.
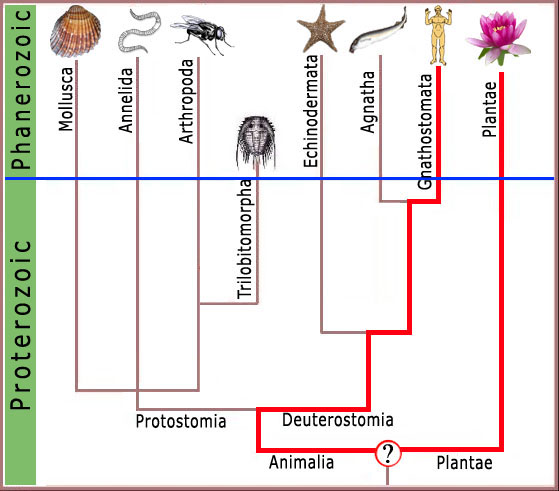
Figura 2. Um quadro representando o tempo geológico padrão no eixo Y, e mostrando a suposta escala da presença de vários grupos de animais e plantas no eixo X. A linha azul fina representa a divisão entre dados (acima da linha) e especulação na ausência de dados (abaixo da linha). Note que qualquer característica biológica molecular comum entre plantas e animais precisam necessariamente ter estado presentes no último ancestral comum de plantas e animais.
Consequentemente, qualquer característica complexa compartilhada pelos artrópodes modernos e pelo homem (Fig. 3), ou pelos artrópodes e pela ervilha (Fig. 4), deveria ter estado presente no ancestral comum a ambas as formas.
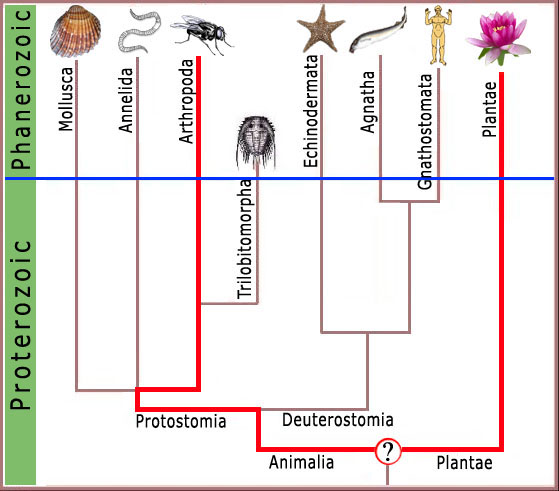
Figura 3. O último ancestral comum entre plantas e artrópodes teria de haver existido pra dentro do Pré-Cambriano, onde não se supõe encontrar nenhuma característica complexa a nível molecular.
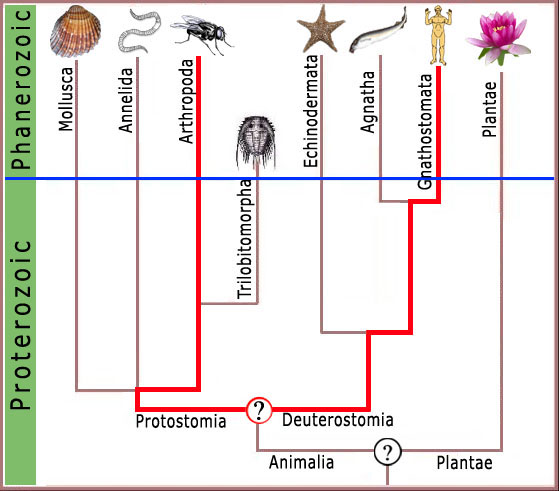
Figura 4. Igualmente entre humanos e artrópodes, o último ancestral comum seria uma bolha desforme e sem características no interior do Pré-Cambriano sem as características complexas dos organismos modernos.
Assim, a presença de características biológicas celulares ou moleculares em comum, entre os artrópodes modernos e o homem ou outras formas de vida, requer que estas características fossem compartilhadas pelo ancestral comum de artrópodes e do homem. Como os trilobitas eram artrópodes, eles também devem ter apresentado estas características e então podemos, confiantemente, atribuir estas características complexas aos primeiros metazoários.
Dentre um grande número potencial de sistemas biológicos moleculares complexos, nós iremos analisar alguns. Será necessário, naturalmente, incluir algum material de natureza bastante técnica, a fim de demonstrar o nível de complexidade presente nas células. Isto é inevitável, pois esta base é necessária para desenvolver os pontos importantes. Estes detalhes são bem conhecidos pelos biólogos moleculares, mas não é necessário ser um biólogo molecular, nem entender os detalhes da complexidade, para entender o significado dos argumentos. Vou começar com a consideração de alguns dos processos básicos compartilhados pelas células dos metazoários. Então iremos examinar algumas das complexas características dos organismos metazoários, incluindo os trilobitas.
O Cromossomo Eucariota
A célula eucariota, que compreende as existentes em todos os organismos com os quais estamos familiarizados, incluindo o homem, carregam a grande quantidade de informação que possuem codificada na forma de longas moléculas de DNA (de menos de 1 cm até mais de 15 cm). Cada célula somática do corpo humano tem 46 destas moléculas. Todo o DNA de uma única célula humana atingiria aproximadamente dois metros, se o DNA das moléculas de todos os 46 cromossomos fossem colocados ponta a ponta.
Este DNA está alojado dentro de um núcleo de aproximadamente 10 micrômetros. Assim, o comprimento do DNA no núcleo de uma única célula humana é 200.000 vezes maior que o raio do núcleo. Uma ilustração equivalente seria colocar 70 quilômetros de fio de pipa em uma caixa de sapatos! Como uma célula consegue fazer isto? Para que ela possa se dividir, precisa primeiramente duplicar totalmente cada cromossomo, produzindo aproximadamente 4 metros de DNA. Depois ela precisa dividir este DNA, precisamente, entre as duas células filhas resultantes. Para apressar este processo, o DNA está separado em cromossomos individuais (Fig. 5), cada um com aproximadamente 50 mm de DNA.
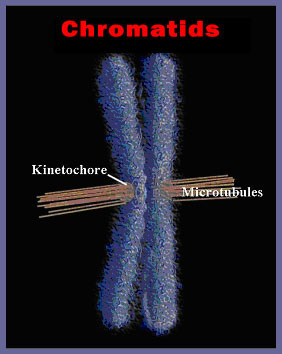
Figura 5. A single human chromosome, here shown doubled just prior to division in the spindle apparatus of the dividing cell. The microtubules have attached to the kinetochore and oriented the chromosome at the equator of the cell. Each of the daughter chromatids contains up to 15 cm (6 inches) of DNA.
Mas este valor é ainda 5.000 vezes maior que o núcleo (figure 6).
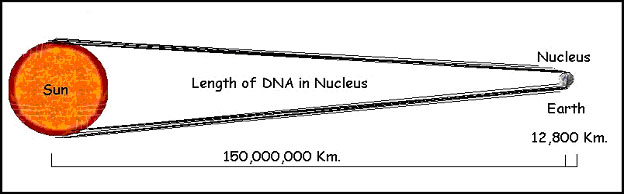
Figura 6. Each cell in our bodies contains approximately 2 meters (6 feet) of DNA. If the nucleus of the cell were the size of the earth, that much DNA would extend to the sun and back eight times!
Portanto, o DNA precisa se organizar de uma maneira muito precisa, para permitir que a célula tenha acesso aos genes necessários e, ao mesmo tempo, permitir que o DNA seja duplicado e dividido com precisão entre as células filhas, durante a divisão celular. Este processo é facilitado, no nível mais básico, pela associação do DNA com uma classe de proteínas denominadas histonas. Estas proteínas muito precisas existem em 5 formas diferentes, conhecidas como H1, H2a, H2b, H3 e H4. As histonas H1, H2a, H2b, H3 e H4, auxiliadas por outras proteínas associadas, formam um octâmero muito estável, contendo 2 cópias de cada molécula. Devido ao fato de todas as histonas terem carga positiva, para permitir que interajam com o DNA, que tem carga negativa, a montagem do octâmero requer a ajuda de diversas proteínas de apoio.
A estrutura do grupo de histonas é tão fundamental para a célula eucariótica que ela é preservada através de todo o espectro de células eucariotas viventes, quase sem modificações. Por exemplo, existem apenas três mudanças de aminoácidos que distinguem a histona H3 da ervilha da do ouriço-do-mar ou do trilobita. A histona H4 humana difere da ervilha por apenas dois aminoácidos! Esta semelhança apresenta outras restrições no problema das origens. Se duas proteínas são tão similares ao longo do espectro inteiro de organismos vivos, devem necessariamente existir severas restrições nas substituições permitidas. Mas evidência experimental sugere que este não é o caso, e que substituições viáveis podem ser feitas, a mais ou menos a mesma taxa, como em qualquer outra proteína de tamanho semelhante. Isto torna essencial a pergunta com relação ao significado das semelhanças, do tempo e dos relógios biológicos. Que mecanismo pode explicar sua origem?
Uma volta e meia de uma molécula de DNA (aproximadamente 140 pares de bases) são então enrolados em torno de cada grupo de histonas, formando um nucleossoma (Figura 7).
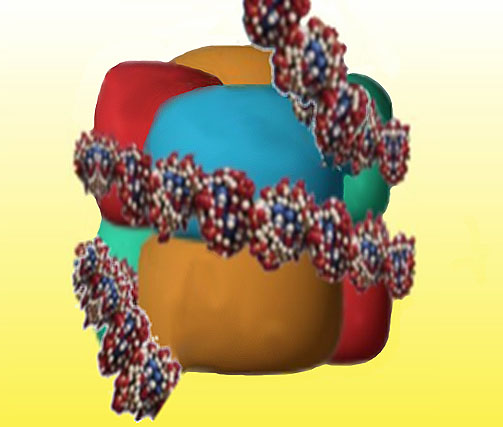
Figure 7. Each Eight of the histone proteins form a cluster that wraps one and a half turns of DNA, greatly reducing the length of the chromosome.
Os nuclessomas são associados em estruturas maiores pela ligação com a histona H1. Estas estruturas, denominadas solenóides, consistem em um arranjo de seis nucleossomas em uma hélice achatada, diminuindo assim a molécula como um todo. Estes solenóides helicoidais são então ancorado ao suporte principal do próprio cromossomo. O suporte principal é composto por uma classe de proteínas, do tipo topoisomerases, que apresentam propriedades extraordinárias. Estas topoisomerases (topo II) estão conectadas à molécula de DNA em sítios específicos. A enzima pode cortar uma das fitas da molécula de DNA no ponto de ligação, agarrar-se às extremidades cortadas, enquanto passa a fita inteira através das extremidades cortadas, ligando então as duas extremidades da fita cortada novamente! A estrutura resultante realizou o inescrutável: condensou uma molécula de DNA de 10 cm de comprimento em uma estrutura 50.000 vezes menor. Mas a complexidade apenas começou.
Cada célula humana tem 46 destas estruturas, que precisam ser duplicadas (92) e então corretamente separadas, de modo que cada célula filha receba um conjunto completo de 46 cromossomos. Noventa e dois corpúsculos separados estão se movendo no citoplasma, em uma viagem infalível para a célula filha correta. Os cromossomos contém um fragmento especial de proteína denominado cinetócoro. A ligação dos microtúbulos à região de ligação do cinetócoro, na cromatina, ocorre quando um microtúbulo, envolvido em uma série de empurrões produzidos pelo rápido alongamento, faz contato com o cinetócoro de uma cromátide e se liga a ele. Se o microtúbulo não fizer contato com o cinetócoro, ele se condensa e então é empurrado em uma direção diferente, até que ele se ligue a um cinetócoro. Quando um número suficiente de microtúbulos, vindos das extremidades opostas da célula, se ligaram aos dois cinetócoros de cada par de cromossomos, os microtúbulos começam a puxar em direções opostas, resultando no alinhamento equatorial dos cromossomos, tão familiar na metáfase.
As duas cromátides se separam no centrômero e são puxadas através do citoplasma, até as extremidades opostas da célula em divisão. O mecanismo de movimento parece ser a contração, expansão e despolimerização dos microtúbulos, que puxam os cromossomos através do citoplasma na direção correta. Estes mecanismos estão presentes em todas as células eucariotas e o envolvimento dos microtúbulos e de proteínas semelhantes à actina e miosina no processo de divisão celular, ilumina a complexidade de um característica que precisa existir em todas as células eucarióticas, incluindo as dos trilobitas, os primeiros fósseis de metazoários registrados. Mantenha isto em mente enquanto exploramos uma característica adicional das células animais em particular: a transmissão do impulso nervoso.
Síntesis de Proteínas orientada:
Síntesis de proteínas é um assunto de inspirar admiração, mas um nós não devemos considerar aqui. Melhor ainda é considerar síntesis de proteínas como dado por um momento, vamos examinar como proteínas são designadas para chegar a seu correto destino na célula por direções contidas dentro de moléculas proteicas.
Muitas proteínas funcionam dentro do citoplasma donde elas são produzidas. Estas proteínas provavelmente necessitam pouca informação para direcioná-las a localizações específicas. Mas um múmero grande de proteínas devem obrigatoriamente chegar a destinos específicos dentro ou fora da célula para serem capazes de funcionar. Por exemplo, algumas proteínas são designadas a funcionar dentro da membrana do retículo endoplasmático da célula. Outras devem obrigatoriamente ser secretadas para fora da célula, ou talvez na membrana mais externa ou interna da mitocôndria, o espaço intramembranoso, ou dentro da matrix mitocondrial. Direcionamento correto da proteína para cada um destas áreas de espaços diferentes requer instruções explícitas dentro da proteína marcada.
No caso da mitocôndria, uma organela de uma célula responsável por conversão de energia guardade em ATP, existem quatro áreas de alvo distintas. Mesmo que a mitocôndria tenha seu próprio DNA e equipamento para síntesis de proteínas, a maioria das proteínas da mitocôndria são feitas de DNA contido no núcleo da célula. Estas proteínas são produzidas no citoplasma, e devem obrigatoriamente navegar de lá para dentro de um compartimento correto da mitocôndria (figure 8).
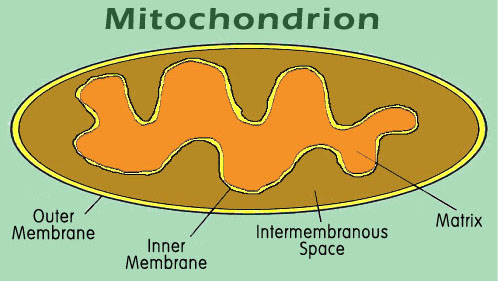
Figura 8. The mitochondrion contains four different target areas that must be separately coded in the protein. The matrix is the site of most of the metabolic activity, and of most of the proteins of the mitochondrion. Both inner and outer membranes are separately targeted by a variety of other proteins, and the intermembrane space is the site for several of the cytochromes.
Os processos de direcionamento são complexos, e alguns dos detalhes estão ainda sendo elaborados, mas muitas das características são bem entendidas. Por exemplo, uma rota possível para uma proteína direcionada a matrix mitocondrial é ilustrada na Figura 9. Cada compartimento da mitocôndria é entregue por um tipo diferente de sinais e receptores sinalares, com o resultado que cada proteína chegue ao seu correto destino.
Algumas proteínas devem permanecer obrigatoriamente dentro da membrana, do retículo endoplasmático, ou dentro da membrana celular mais externa, ou em outra membrana celular. Certo tipo de proteína desenvolve uma função vital na regulação da passagem de materiais através da membrana, e em outros processos celulares vitais. Para que a proteína seja produzida nesta configuração, o gene para sua produção deve obrigatoriamente conter, em adição a informação usual sobre como construir uma proteína funcional ativa, uma variedade de instruções informando a célula qual o destino e caminho que a proteína deve seguir.
Um dos possíveis destinos para uma proteína é a membrana celular mais externa. Proteínas destinadas para este propósito começam com um plano especial de instruções chamado um sinal. Este sistema de sinais é reconhecido por um corpo citoplasmático especial chamado de partícula de reconhecimento de sinais (SRP) (figure 9).
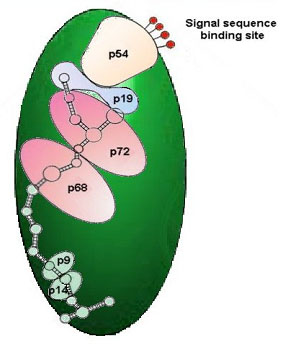
Figura 9. The Signal-Recognition Particle (SRP) is part of a complex of proteins responsible for targeting proteins to specific compartments of the cell. The mechanisms, components and the targeting information appear to be universal, being recognized in plant, animal, and even yeast cells. Proteins destined for targets outside of the cytoplasm (either in membrane-bound compartments, or in the membranes themselves, or for secretion outside the cell), are designated by specific sequences of amino acids in the leader region of the protein. The particle responsible for identifying these specific sequences, called signal peptides in nascent (growing) proteins, is the SRP. This complex consists of a chain of 300 specific bases of RNA and six proteins, identified by their respective molecular weights (in kilodaltons): P9, P14, P54, P68 and P72. It is known that the P54 protein is responsible for reading and interacting with the signal peptide, the two small proteins interact with the ribosome, and the large P68/P72 proteins are involved in the movement of the nascent peptide chain. The SRP will stop protein synthesis after about 70 amino acid residues, in the absence of suitable membrane interactions, preventing the synthesis of proteins in inappropriate environments.
Esta partícula identifica uma mensagem em código nos primeiros 50 ou mais de aminoácidos das proteinas não citoplasmáticas a medida que elas são produzidas pelo ribossomo, e conecta com esta sequência de liderança, referrida como o sinal do peptídeo. Os passos subsequentes são detalhados em uma serie de diagramas.
Moléculas como as de proteinas com canais de sódio com voltagems juntas discutidas abaixo devem ter sido obrigatoriamente codificadas dentro de todas elas com a informação para seus atributos funcionais e estruturais, e com a informação para adquirir suas funções ativas distribuídas através de todo o comprimento da molécula.
Este mecanismo de sinais é universal, desde que os processos operam no mesmo caminho virtualmente em todas as células eucarióticas. Em adição, as proteínas comprimem o translocon e o receptor SRP, que são responsáveis pela inserção de membranas direcionadas de proteínas dentro da membrana também são proteínas de membranas fechadas. Isto significa que elas também devem ser inseridas dentro da membrana por um mecanismo similar. A universalidade significa as proteínas e o mecanismo por suas aquisições por membranas os quais estavam bem antes presentes nos primeiros recordes de metazoa, e também na célula eucariótica ancestral.
A Sinapse:
A célula nervosa ou neurônio em repouso tem um potencial elétrico de membrana de 60 milivolts negativo, no interior da célula. Este potencial é estabelecido por uma bomba especial de sódio/potássio, que utiliza a energia celular para bombear íons sódio positivamente carregados para fora da célula. O impulso nervosa é iniciado e propagado por um influxo de íons sódio para o interior da célula, através de canais de sódio, formados por uma proteína especial, e que se localizam na membrana celular. A propagação é mediada pela abertura sucessiva dos canais de sódio da membrana, ao longo de todo o axônio. Esta proteína passa através da membrana celular externa 24 vezes (figura 10).
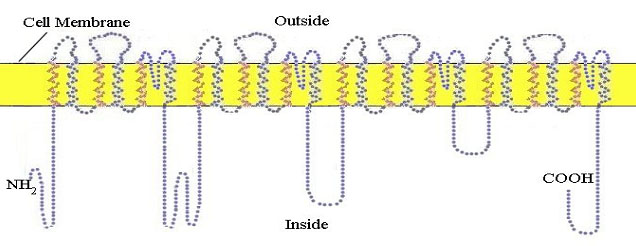
Figura 10. The 24-pass voltage-gated sodium channel protein shown here at completion of its synthesis in the membrane of the endoplasmic reticulum prior to incorporation into the outer cell membrane.
As proteínas que compõem o canal de sódio operado por voltagem são não somente muito complexas em sua construção, mas são planejadas de forma intrincada para exercer seu papel de forma surpreendente. A proteína forma um canal através da membrana exterior da célula que contém um portão operado por voltagem e um guarda de fluxo reverso crítico (figura 11).
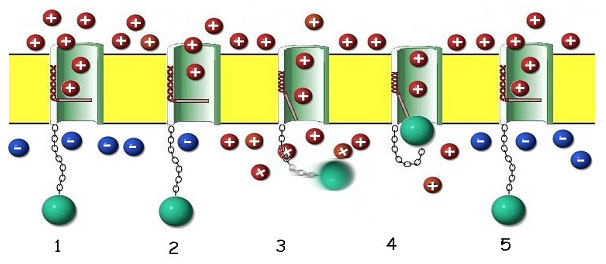
Figura 11. The 24-pass Voltage Gated Sodium Channel Protein as it may appear in its assembled form within the outer cell membrane (yellow). 1. When the membrane is at rest, the channel is blocked by a gating portion of the protein. The resting membrane maintains a positive charge on the outside and a negative charge on the inside. This charge disparity accrues as a consequence of two factors. A sodium-potassium pump (not shown here) forces sodium (red, positive charge) outside the cell membrane, leaving the anionic (blue, negative charge) proteins behind in the cytoplasm. 2. When the sodium channel protein senses a change in voltage resulting from the opening of adjacent pores (depolarization), a solenoid-like portion of the protein in the wall of the channel (red coil) begins to contract. 3. The coil opens the gate as it contracts, allowing sodium ions to rush through the channel into the cell, motivated by diffusion and the potential gradient. The timing of this process is critical. If the channel continues to admit sodium, the cell cannot respond to further stimuli and would quickly die. 4. Since channel cannot close while the membrane is depolarized, a special portion of the protein (green ball) acts like a plug to close the channel until the sodium-potassium pump can restore the resting membrane potential. 5. In a few milliseconds, the pump has removed the excess sodium, the membrane potential is restored, the gate is closed, and the plug relaxes, ready for the next stimulus.
Quando a despolarização do nervo é sentida pela proteína porina, o portão se abre e íons sódio fluem para dentro da célula, propagando a mudança de potencial e provocando a mesma resposta nos póros adjacentes. Quando a membrana fica completamente despolarizada, um segmento bloqueador especial tampa o canal, evitando mais despolarização, até que o potencial de membrana em repouso tenha sido restabelecido pela bomba de íons sódio. Quando o impulso nervoso atinge a extremidade de um nervo, ele precisa passar o sinal através de um espaço para a próxima célula nervosa. A junção das duas células é chamada sinapse, e o espaço separando as duas células de fenda sináptica (figura 12).
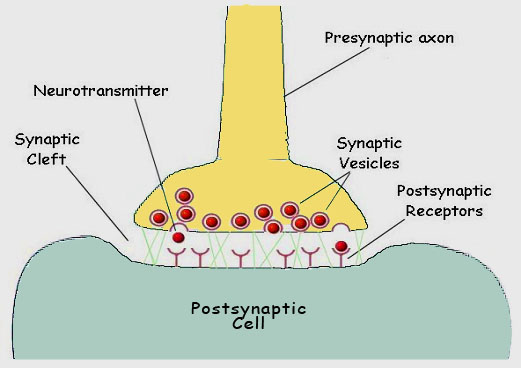
Figura 12. The chemical synapse. When one nerve must pass an impulse on to the next cell, it does so by one of two general processes. Where speed is the critical factor, a direct electrical connection may exist between the two cells Where modulation is important, a chemical event intercedes between the arrival of the impulse at the terminal of one axon and the continuation of the signal in the next cell. The juncture is referred to as a synapse. The cells are separated by a gap, the synaptic cleft. Upon appropriate stimulation the presynaptic cell releases neurotransmitter (commonly acetylcholine). The neurotransmitter then diffuses across the cleft to receptors on the postsynaptic membrane. Here the receptors respond by initiating depolarization of the membrane and the impulse is propagated in the postsynaptic cell.
Em muitas células, a transmissão do impulso nervoso é mediada pela liberação de uma substância neurotransmissora, geralmente a acetilcolina, que é uma molécula pequena. A acetilcolina é acumulada em vesículas membranosas citoplasmáticas, onde uma proteína "antiport" de íons hidrogênio troca a acetilcolina, produzida no citoplasma da célula, por íons hidrogênio, que são bombeados para dentro da vesícula às custas de energia obtida por hidrólise do ATP. A vesícula é então transportada pelo citoplasma através de microtúbulos do citoesqueleto até a membrana da superfície sináptica (figura 13).
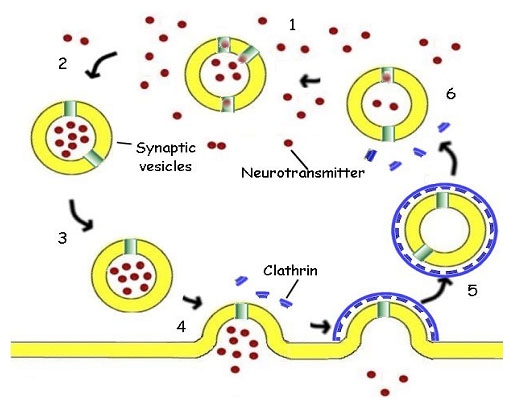
Figura 13. The Synaptic Vesicle. 1. The synaptic vesicle accumulates hydrogen ions at the expense of ATP. The hydrogen ions are then exchanged for acetylcholine (red), a neurotransmitter, by a specific antiport protein. 2. As the vesicle becomes charged with acetylcholine, it is picked up by a cytoplasmic transport protein and carried through the cytoplasm to the synaptic region of the neuron. 3. The filled vesicle is docked in the region of the synapse, where it awaits a nerve stimulation. 4. Triggered by an influx of calcium ions, the vesicle releases its charge of acetylcholine to the synapse, passing the impulse on to the post-synaptic cell. 5. The empty vesicular membrane is surrounded by cytoplasmic protein molecules called clathrin (blue), capturing the vesicle for reuse. The clathrin forms a cage coating the entire vesicle. The vesicle then travels back away from the membrane into the cytoplasm and loses its protective cage. 6. The vesicle again begins to accumulate hydrogen ions the cycle is repeated.
Este processo é surpreendente por si mesmo, porque moléculas do tipo quinesina simplesmente andam sobre os elementos do citoesqueleto de uma maneira bastante antropomórfica, carregando a vesícula sináptica com elas.
Na membrana da vesícula existem algumas proteínas singulares, que não são encontradas em nenhum outro lugar de membrana celular. Entre elas estão a sinaptobrevina e a sinaptotagmina. A sinaptobrevina se liga a um complexo de proteínas chamado NSF (Fator Sensível à N-etilmaleimida) e SNAPs (proteínas associadas ao NSF solúvel). Este complexo se liga à sintaxina, uma proteína encontrada apenas na membrana plasmática, na região da sinapse, ancorando assim a vesícula na membrana. A sinaptotagmina, a outra proteína vesicular mencionada, tem dois sítios de ligação para íons Ca++ do seu lado citoplasmático. Na ausência de Ca++, a sinaptotagmina se liga ao complexo sinaptobrevina– sintaxina–NSF–SNAP e impede a ligação da alfa SNAP, a proteína de fusão. Quando o impulso nervoso chega na região da sinapse, abrem-se os canais de cálcio, permitindo a entrada de Ca++ no citoplasma. A sinaptotagmina se liga ao cálcio e então a alfa SNAP pode se ligar ao complexo. Como conseqüência, a membrana da vesícula se funde com a membrana celular, por um mecanismo ainda não bem compreendido, expelindo o conteúdo da vesícula na sinapse, e provocando a resposta da célula vizinha.
Outras proteínas que também estão envolvidas na separação e dispersão das vesículas citoplasmáticas são as proteínas Rab, que constituem o sistema UPS da célula. Estas proteínas se ligam a todas as vesículas no citoplasma, funcionando como "rótulos de expedição", para especificar o seu destino final. Quando elas chegam ao seu destino, o "rótulo de expedição" é lido e, se o destino é aquele especificado, permite-se que as vesículas se fundam e compartilhem os seus conteúdos com a organela receptora. Se o rótulo especifica algum outro destino, o acesso da vesícula à organela é recusado.
Enquanto isso, uma proteína citoplasmática denominada claterina identifica a vesícula vazia e a cerca com uma gaiola hexamérica, que preserva a membrana e evita que as proteínas associadas sejam perdidas. A gaiola de claterina permanece unida à vesícula até que esta possa se reunir com o seu endossoma hospedeiro no citoplasma, para se encher novamente. Este processo, que eu descrevi de maneira bastante simplificada, é comum em todos os animais que possuem sistema nervoso, dos mais simples invertebrados ao homem. Devido ao fato deste processo representar um mecanismo muito complexo, compartilhado por insetos e homens, podemos estar absolutamente certos de que ele existia também nos trilobitas.
A Biologia do Desenvolvimento dos Trilobitas
O que podemos dizer sobre as vias metabólicas complicadas pelas quais um simples óvulo, no ovário de uma mãe trilobita, se transforma num descendente viável? Muito mais do que você poderia imaginar, graças aos avanços recentes em nossa compreensão da biologia molecular do desenvolvimento. Aqui eu só poderei dar um esboço dos detalhes. Deixe-me rapidamente contar um pouco sobre como um inseto é formado. Aqui nós vamos discutir sobre a mosca das frutas, Drosophila, que é um inseto que sofre metamorfose. Como estes insetos são bastante pequenos, não seria prática a eclosão de um descendente com asas, completamente funcionais, a partir de um óvulo fertilizado. A estratégia de muitos insetos é colocar um ovo, o qual "eclode" originando um ovo maior, chamado de larva. A larva é apenas um saco para acumular material alimentar, na preparação para a produção da forma adulta. Entretanto, nos recônditos mais profundos de cada larva, estão as sementes embrionárias de um organismo adulto. Denominados "discos imaginários", estes tecidos especializados permanecem dormentes até a formação da pupa. Neste momento a larva se dissolve e os discos imaginários se transformam nas várias partes do adulto. Este é um processo surpreendente por si só, mas a seqüência de eventos que levam à formação dos discos imaginários oferece uma visão sem precedentes do processo de desenvolvimento, que nos será de grande interesse em nossa consideração sobre o trilobita.
Enquanto o ovo ainda está dentro do ovário, são estabelecidos gradientes específicos de produtos gênicos reguladores dentro do ovo. Estes m-RNAs ou proteínas se originam do próprio núcleo do ovo ou de células maternas acessórias que rodeiam o ovo no ovário. Após a fertilização, são ativadas séries adicionais de genes, produzindo proteínas reguladoras adicionais, em regiões específicas do ovo fertilizado. Esta distribuição assimétrica de proteínas reguladoras resulta em cada célula tendo uma combinação única de reguladores. O balanço desses genes reguladores determina quais genes são ativados e quais são suprimidos em cada célula, e esta assimetria, por sua vez, determina cabeça e cauda, e diferenciação ao longo dos eixos do corpo resultantes.
Este sistema de desenvolvimento é fantasticamente complexo. Estudos genéticos em Drosophila revelaram uma classe de genes do desenvolvimento que, quando sofriam mutação, resultavam não apenas em uma única alteração, tal como a cor do olho, mas produziam efeitos em massa que eram letais, ou resultavam em mudanças monstruosas na forma do corpo. Por exemplo, uma única mutação gênica em um dos genes reguladores resultou no crescimento de pernas no local onde se encontram normalmente as antenas, ou na formação de um segmento extra no corpo, com um par extra de asas. Vastas redes reguladoras ligam cada um desses genes do desenvolvimento a centenas de outros genes.
Para seu assombro, investigadores descobriram que os genes que estavam controlando o desenvolvimento das moscas das frutas e os genes que controlam o desenvolvimento dos vertebrados, incluindo camundongos e homens, tinham estrutura muito semelhante e freqüentemente controlavam partes análogas dos embriões das moscas e do homem. E assim, estas seqüências de genes do desenvolvimento, presentes nas moscas e no homem, também estavam presentes nos trilobitas.
Estudos subsequentes revelavam a localização de alguns destes genes no cromossomo. Quando se identificou e mapeou a série principal de genes reguladores que determina a polaridade do embrião de Drosophila (genes HOM-C), os investigadores descobriam um fato surpreendente, um fato que eles não esperavam e não estavam preparados para lidar com ele quando o encontraram: os genes que controlavam o desenvolvimento do eixo do embrião, da cabeça à cauda, estavam localizados no cromossomo na mesma ordem que a das porções da anatomia do organismo cujo desenvolvimento eles controlavam (colinearidade). Isto é inesperado por muitas razões, sendo uma delas a improbabilidade de tal arranjo ocorrer na ausência de um projetista.
Alguns anos atrás, Murry Eden, um matemático do M.I.T., demonstrou a improbabilidade de se obter genes com uma seqüência específica no cromossomo. Parece não haver nenhuma razão funcional para estes genes estarem tão organizados, apesar de que este quadro pode mudar. Mas este não era o fato mais assombroso. Estudos subsequentes em vertebrados (principalmente nos camundongos, mas também no homem) mostraram que tipos semelhantes de proteínas reguladoras eram responsáveis pelo ordenamento da organização da cabeça até a cauda do corpo dos vertebrados, incluindo o homem. E estes genes (chamados de genes Hox), que eram muito semelhantes aos genes equivalentes na Drosophila (em alguns genes homeóticos, a similaridade entre Drosophila e o homem é de 98%) estão localizados no cromossomo na mesma ordem que a das moscas das frutas!
Eles devem ter tido uma origem comum! E eles deviam estar presentes no trilobita, a primeira forma metazoária do cambriano. Assim, nas primeiras formas, não só estavam presentes todas as complexidades da célula eucariota, mas também estava presente toda a complexidade insondável do sistema de desenvolvimento, envolvendo a interação de milhares de genes, que todas as formas cefalizadas parecem ter em comum.
O Olho do Trilobita
O olho tem sido um objeto de assombro durante toda a história registrada, devido às suas funções críticas. Certamente a existência de olhos compostos e completamente funcionais nos primeiros metazoários tem feito com que, de quando em quando, evolucionistas pensantes questionem seriamente sobre a base de sua origem. No caso dos trilobitas, não só as primeiras formas a aparecem estavam equipadas com órgãos visuais altamente organizados, mas algumas das propriedades recentemente descobertas dos olhos dos trilobitas representam uma "proeza em otimização de função". Os olhos dos trilobitas (figura 14),

Figura 14. A trilobite eye illustrating the complex compound structure.
a partir do que podemos concluir pelo estudo das formas fósseis, têm muito em comum com os olhos dos insetos modernos (figura 15).

Figura 15. . The eye of a modern insect illustrating a compound structure similar to that of the trilobite.
Certos trilobitas de meados do Paleozóico têm um sistema ótico único, desconhecido em qualquer outra criatura. O físico nuclear Levi-Setti (Diretor do Fermilab da Universidade de Chicago), que é aficcionado por trilobitas, afirma com franqueza impertubável:
"E uma descoberta final – a de que a interface refratora entre os dois elementos das lentes do olho do trilobita foi projetada [ênfase adicionada] de acordo com as construções óticas elaboradas por Descartes e Huygens em meados do século dezessete – toca as raias da pura ficção científica."
Os eixos dos omatídeos individuais foram construídos a partir de cristais únicos de calcita, sendo o eixo ótico do cristal coincidente com o eixo ótico do elemento do olho. Isso representa um problema extraordinário para o trilobita, já que uma simples lente esférica e grossa de calcita não poderia decompor a luz em uma imagem (figure 16).
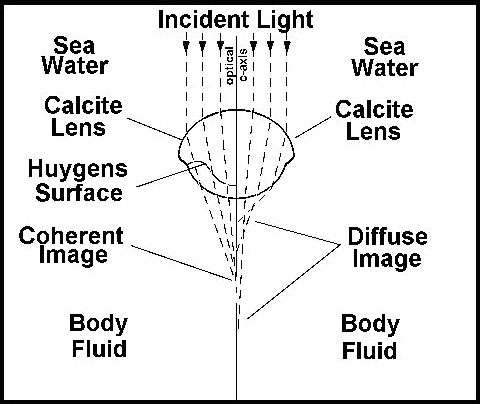
O elemento ótico do trilobita é uma lente composta, constituída por duas lentes com diferentes índices de refração, unidas em uma superfície de Huygens. Para que tal olho focalize a luz corretamente nos receptores, ele teria que ter exatamente esta forma de lente. Os princípios óticos requeridos foram elaborados pela primeira vez por Huygens no século dezessete, mas as lentes dos trilobitas funcionavam perfeitamente, usando estes princípios óticos, muito antes desse matemático holandês entender como. Os primeiros trilobitas não tinham estas lentes sofisticadas, mas os seus olhos eram, aparentemente, mais semelhantes aos dos insetos modernos. Porém, não se conhecem formas intermediárias no registro fóssil. Quando a lente de Huygens apareceu pela primeira vez, ela era completamente funcional.
O mecanismo regulador do desenvolvimento do olho deve realmente ser complexo. Estima-se que 2.500 a 5.000 genes estão envolvidos no processo de desenvolvimento. Alguns dos detalhes do desenvolvimento estão sendo solucionados em Drosophila, onde alguns dos genes de ligação mestres são conhecidos. Cada faceta individual ou omatídeo de um olho composto, tal como o da Drosophila, consiste de um grupo de oito células, sete das quais se desenvolverão em receptores de luz. Descobriu-se que uma dessas células retinais, chamada R7, era responsável pela detecção da luz ultra-violeta. A via do desenvolvimento da R7 tem sido assunto de uma intensa investigação por muitos anos. Ela revelou uma cascata de interações que parece representar a maioria das vias de ligação externa das células.
A membrana da célula R7 contém uma proteína especial, o receptor da tirosina quinase (RTK). Esta proteína possui um sítio receptor extracelular, uma região transmembrana e uma região enzimática intracelular. Quando algum ligante externo se liga ao receptor (neste caso é o ligante de membrana da oitava célula), a molécula se une com outro RTK, formando um dímero. As duas moléculas então se empenham na fosforilação recíproca de três resíduos específicos de tirosina, cada um na outra molécula.
Assim fosforilada, a região citosólica pode se ligar a uma proteína citoplasmática específica (GRB2) que reconhece o RTK fosforilado. Quando a GRB2 se liga ao RTK, este pode então se ligar a uma terceira proteína, Sos. O complexo Sos faz com que a proteína associada à membrana, Ras, perca GDP, que é então substituído por GTP. Nesta condição, a proteína Ras se liga à proteína chamada Raf, uma quinase da treonina/serina. Quando ligada ao Ras ativado, Raf é capaz de ligar e fosforilar outra quinase específica da tirosina/treonina, a MEK, ativando-a. MEK por sua vez ativa uma enzima citoplasmática, a MAP quinase, através da fosforilação dos resíduos de tirosina e treonina nesta enzima. A MAP quinase está aparentemente envolvida na fosforilação das proteínas de ligação ao DNA e outras proteínas celulares chave, que resultam na mudança da direção da diferenciação celular, de tal modo que tal célula agora se transformará numa R7 normal. O que é especialmente digno de nota nesta cascata, é que ela é encontrada em todas as células de organismos eucariotas multicelulares, e, com pequenas alterações, também nos eucariotas unicelulares ( leveduras e protozoários).
Recentemente, como resultado da manipulação de um gene mestre do desenvolvimento, o "eyeless", produziram-se moscas com olhos em várias partes do corpo, incluindo as asas, pernas e extremidades das antenas, como resultado da ativação do gene em posições anormais. Um gene mestre semelhante foi encontrado nos vertebrados, que possuem olhos completamente diferentes dos insetos. O gene do homem, camundongo e outros animais é quase idêntico ao da Drosophila. Quando o gene apropriado de um cromossomo do camundongo (e possivelmente também de um humano) foi inserido em uma mosca, ele produziu olhos de mosca em todos os lugares do corpo em que foi ativado. Os dois genes são suficientemente semelhantes, de modo que o gene de mamífero leva à formação de um olho de mosca. Nossa linha de raciocínio nos leva a concluir que o sistema de genes que leva ao desenvolvimento do olho estava presente e funcionando nos primeiros trilobitas.
Há um número cada vez maior de vias metabólicas do desenvolvimento que são "conservadas evolutivamente", um eufemismo para "embaraçosamente semelhantes", em um largo espectro de organismos, e a maioria delas estaria presente nos trilobitas. Por exemplo, os genes responsáveis pela organização da dorso-ventralidade no homem foram descobertos, utilizando os genes de Drosophila como sonda molecular. Os genes responsáveis pela organização do cérebro humano na embriogênese foram descobertos, utilizando os genes de Drosophila como sonda. O olho, a parte posterior do cérebro e a medula espinhal, o "pathing" dos axônios, a diferenciação dos músculos do esqueleto e do coração, a resposta fotoperiódica, a escultura dos tecidos envolvendo a morte de células selecionadas (apoptose), a modelagem embrionária, a sinalização celular e milhares de outros exemplos de processos "conservados evolutivamente" poderiam ser citados. Até a formação dos membros é dirigida, nas moscas das frutas, por um gene (Hedgehog), cujo gene homólogo nos vertebrados (Hedgehog Sônico) comanda a formação dos membros em todos os vertebrados conhecidos, incluindo o homem, camundongo, galinha e até o peixe! O mecanismo elaborado responsável precede claramente qualquer organismo com membros conhecido. De onde veio toda essa informação?
O Problema da Complexidade dos Primeiros Metazoários
Eu só apresentei uns poucos exemplos breves ilustrando a complexidade das células e organismos eucariotas atuais. Estes foram extraídos dentre centenas ou milhares de outros exemplos que poderiam igualmente ter sido usados para lograr o intento. Os primeiros animais complexos dos quais se tem um registro conhecido, os trilobitas, apareceram no início do Cambriano. A base do Cambriano é, algumas vezes, definida vagamente como o ponto na coluna geológica onde aparecem os primeiros trilobitas.
Os trilobitas são artrópodes, o mesmo grupo dos insetos modernos. As células dos trilobitas se dividiam de maneira semelhante a de todos os eucariotas modernos. Os mecanismo estavam todos no lugar, todos funcionando como hoje. Os trilobitas tinham sistemas nervosos tão complexos como os dos insetos modernos. As sinapses do sistema nervoso dos trilobitas funcionavam como as sinapses de todos os organismos modernos. Os olhos dos trilobitas manifestam toda a complexidade e integridade de desenvolvimento das formas modernas. As complexidades que acabamos de descrever estavam todas presentes, todas funcionando completamente nos PRIMEIROS ANIMAIS MULTICELULARES DOS QUAIS TEMOS REGISTROS. ONDE E QUANDO OCORREU A EVOLUÇÃO???
Esta questão tem sido habilmente evitada pelos evolucionistas. Os sistemas que acabamos de descrever não aconteceram por acaso. Eles foram planejados. Cada passo dado por um trilobita foi uma acusação das impropriedades da teoria da evolução. Isto explica porque, quando evolucionistas como Steven Gould escrevem livros sobre as primeiras formas de vida, eles cuidadosamente evitam mencionar o problema do aparecimento súbito de formas infinitamente complexas. Suas atitudes são " Estas coisas estão lá, portanto a evolução deve ser capaz de fazê-las". A evolução é uma caixa preta. Ela é mágica! Isto é EVOLUÇÃO!
Em relação à escassez de evidência fóssil no Pré-cambriano, Leonard Brand (1998) escreveu:
"Uma de suas [cientistas evolucionistas] maiores suposições foi a de que o relógio molecular é confiável. Quando Levington apresentou seu trabalho [nas reuniões do GSA de 1996, em Nova Orleans] ele afirmou que o relógio molecular pode ser melhor comparado com um relógio de sol na sombra, o que não é muito encorajador para o seu método, mas ele e seus colaboradores ainda acreditam que este relógio produziu dados suficientes para testar a teoria da evolução rápida da vida no início do Cambriano... .
A partir dos dados do relógio molecular eles concluíram que a divergência inicial das formas de vida metazoárias ocorreu há, aproximadamente, 1,2 bilhões de anos (mais ou menos 50 a 250 milhões de anos). A base do Cambriano é considerada geralmente como 543 milhões de anos atrás. Assim, suas conclusões requerem meio bilhão de anos de vida metazoária antes do Cambriano. Eles também concluíram que o início dos filhos metazoários não foi uma explosão, mas sim algo bastante espalhado durante aquele meio bilhão de anos.
Alguns dias mais tarde estes trabalhos foram discutidos na "Discussão de temas quentes" na hora do almoço. Quatro cientistas fizeram apresentação breves sobre novas idéias concernentes à explosão Cambriana, seguidas por perguntas e comentários da audiência. Muitas perguntas foram sobre detalhes do método de pesquisa utilizado, mas duas questões se destacaram. É necessária uma pequena explicação antes de lidar com estas questões. A proposta de que animais metazoários complexo, ancestrais de seres como moluscos, trilobitas, vertebrados, ouriços-do-mar, corais e muitos outros, existiram durante meio bilhão de anos antes do Cambriano, implica que eles viveram todo aquele tempo sem deixar registro fóssil. Isto requer que, antes do Cambriano, eles tenham existido como animais em forma de vermes moles ou semelhantes a larvas, com a constituição genética geral dos grupos cambrianos, mas sem sua morfologia com esqueletos.
Agora as questões. A primeira das duas foi – Por que os fósseis de vestígios (rastros, trilhas e tocas fósseis) são tão raros da base do Cambriano, se estes animais existiram durante meio bilhão de ano? Um especialista em fósseis de vestígios, reconhecido internacionalmente, se levantou, possivelmente para responder a pergunta. Entretanto, ele falou sobre outras coisas e a questão mais importante nunca foi respondida. No final da discussão, outro cientista se levantou e comentou sobre a implicação de que todos os filos que possuem esqueletos desenvolveram-nos mais ou menos ao mesmo tempo no Cambriano. Ele perguntou: - Por que todos esses tipos de animais estavam vivendo há tanto tempo e então todos resolveram produzir esqueletos ao mesmo tempo? Ele então perguntou, com certo vigor – "Por que vocês estão evitando a pergunta verdadeira?" Depois de uma pausa, um dos apresentadores respondeu "Porque é realmente difícil (uma pergunta difícil)". Ele continuou dizendo que eles esperavam que as respostas surgissem após estudo mais profundo da biologia do desenvolvimento.
Estas duas perguntas não foram feitas, aparentemente, por pessoas que duvidavam da teoria da evolução, mas por cientistas evolucionistas desejosos de fazer as perguntas difíceis que precisam ser formuladas enquanto eles tentam testar diferentes hipóteses. Permanece o fato de que a explosão no Cambriano é um dos maiores desafios para as teorias naturalistas, que ainda continua sem respostas".
Algumas citações recentes de um artigo na revista Time (When life exploded, TIME, 12/05/95), sobre a origem dos metazoários pré-cambrianos, são úteis em nos ajudar a compreender o quão perto da verdade alguns evolucionistas estão chegando. O artigo salienta que todos os filos animais, exceto talvez os briozoários, estavam presentes no início do Cambriano e que todos eles apareceram em um período de tempo muito curto ("não mais que 10 milhões de anos").
Steven Gould, da Universidade de Harvard (paleontólogo) "Rápido é agora muito mais rápido do que pensávamos, e isto é extraordinariamente interessante".
Samuel a Bouring, M.I.T. (geólogo): "Nós agora sabemos quão rápido o rápido é e o que eu gostaria de perguntar para os meus amigos biólogos é: Quão rápida deve se tornar a evolução antes que comecemos a nos sentir desconfortáveis?"
Rudolph Raff, Universidade de Indiana (biólogo): "Devem existir limites para a mudança. Afinal, nós temos estes mesmos velhos planos corporais há meio bilhão de anos".
G.M. Narbone, Universidade de Queens (paleontólogo): "O que Darwin descreveu no livro Origem das Espécies era o tipo de evolução com base estável. Mas também parece haver um tipo de evolução não-darviniana que atua em períodos de tempo extremamente curtos – e é lá que está toda a ação".
Conclusão:
Fizemos uma consideração cuidadosa de uma pequena amostragem dos milhares de exemplos de complexidades compartilhadas pelas formas modernas, que poderiam ter sido utilizados. Vimos que, a partir de uma cuidadosa consideração da evidência, a teoria da evolução não explica a origem dos sistemas ricos em informação dos organismos biológicos.
Vimos que os trilobitas, os primeiros fósseis abundantes e bem representados de metazoários, apresentavam uma complexidade inimaginável em cada detalhe, com olhos compostos, com apêndices abdominais e brânquias, com pernas e antenas e com formas esculturais intrincadas e complexas. Eles possuíam sistemas musculares e nervosos totalmente funcionais. O seus olhos se desenvolviam por processos não só semelhantes aos dos outros artrópodes, mas semelhantes aos dos vertebrados, incluindo o homem. O sistema complexo de desenvolvimento das formas cefalizadas já estava presente e funcionando. Milhares de outras complexidades de biologia molecular, compartilhadas pelas formas modernas, já estavam operando. De onde vieram estas complexidades?
O trilobita, entre os primeiros organismos a existirem na parte mais inferior da coluna geológica, era COMPLETAMENTE complexo a nível molecular tanto quanto qualquer forma moderna de vida. Não há evidência da existência de forma anterior alguma da qual eles poderiam ter derivado. Além disso, não há evidência da existência de algum mecanismo, nos sistemas biológicos, que adicione informações a sistemas complexos (Spetner, 1998). Argumentar que eles vieram de formas pré-cambrianas que não se preservaram porque não tinham partes duras é argumentar novamente sobre a AUSÊNCIA DE EVIDÊNCIA.
A ausência de evidência, em ciência, deve ser interpretada como evidência de ausência. Não existe seqüência evolutiva pré-cambriana porque não houve evolução pré-cambriana. A evolução como uma explicação para a existência de sistemas viventes complexos é um ponto de vista religioso, sustentado por aqueles que desejam que o mundo não tenha nenhum Originador (inferindo sobre - Huxley 1937, p. 312).
Os trilobitas e todas as outras formas apareceram em cena como organismos completamente formados e completamente competentes. Já passou da hora de substituir a teoria da evolução orgânica por uma teoria que possa explicar os dados. A única teoria que pode explicar a origem da informação é a teoria da Criação Especial. Eu não peço desculpas por escolher colocar a minha fé na existência de um Mestre Projetista, uma posição que é consistente com a mais clara interpretação da evidência disponível no registro fóssil, consistente com a mais clara leitura do livro de Gênesis e uma fé que é positiva, enaltecera e cheia de esperança para o futuro.
Bibliografia
Brand, L. 1998. Comunicação pessoal .
Lodish, Harvey, et. al. 1995. Molecular Cell Biology. Third edition. Scientific American Books. W. H. Freeman and Co. New York.
Dobzhansky, Th., F. Ayala, G. L. Stebbins and J. W. Valentine. Evolution. 1977. (W. H. Freeman &Co.San Francisco). p.265, ff.
Wray, G. A., Levinton, J. S. & Shapiro, L. H. 1996, Molecular evidence for deep pre-Cambrian divergences among metazoan phyla. Science, 274, 568-573.
Carroll, Sean. 1995. "When Life Exploded." Time Magazine. 146:(23). Boncinelli, E., Simeone, A., La Volpe, A., Faiella, H., Acampora, D., & Scotto, L. 1985. Human cDNA clones containing homeobox sequences. Cold Spring Harb. Symp. Quant. Biol., 50, 301-306.
Levi-Setti, R. Trilobites. 1993. (University of Chicago Press, Chicago). p. 29. Ibid, p.54.
Rubin, Gerry, Developmental Geneticist U.C. Berkeley. Quoted in "Secrets in a fly’s eye", Discover 17: (7) 110.
Levinton, J. S., G. Wray, and L. Shapiro. 1996. Molecular evidence for a deep Precambrian divergence of animal phyla. I. Introduction and regression approach. Abstracts with Programs, 28 (7): A-52. Geological Society of America annual meeting, Denver, CO.
Wray, G., J. S. Levinton, and L. Shapiro. 1996. Molecular evidence for a deep Precambrian divergence of animal phyla. II. Relative rate tests and implications. Abstracts with Programs, 28 (7): A-52. Geological Society of America annual meeting, Denver, CO.
Grotzinger, J. P., S. A. Bowring, B. Z. Saylor, and A. J. Kaufman. 1995. Biostratigraphic and geochronologic constraints on early animal evolution. Science 270:598-604.
Huxley, Aldous. 1937. Ends and Means.
Spencer, Lee A. 1998. Not by Chance! Judaica Press.